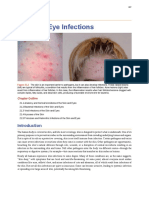
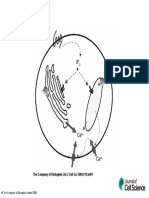
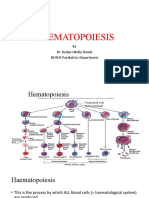
You might also like
- VaccinesDocument39 pagesVaccinesritu9456No ratings yet
- Microbiology NotesDocument3 pagesMicrobiology NotesAthena Huynh100% (1)
- Immunosuppressants - AMBOSS PDFDocument7 pagesImmunosuppressants - AMBOSS PDFOpio IsaacNo ratings yet
- Human Skin 1Document9 pagesHuman Skin 1Baciu DianaNo ratings yet
- Bob Beck Lecture - Take Back Your PowerDocument42 pagesBob Beck Lecture - Take Back Your Powerprobiermalaus100% (1)
- Physicians Approach to Biotoxin IllnessDocument81 pagesPhysicians Approach to Biotoxin IllnessCody ThomasNo ratings yet
- Oet Test Material: Occupational English Test Reading Sub-Test NursingDocument13 pagesOet Test Material: Occupational English Test Reading Sub-Test NursingRyu Tse33% (3)
- Burns Trauma.Document44 pagesBurns Trauma.armaan hussain100% (1)
- Immunology KubyDocument24 pagesImmunology KubySayanta Bera100% (1)
- Chinese Medicine 2Document23 pagesChinese Medicine 2Tim SchlankNo ratings yet
- Treatment of Dry Skin SyndromeDocument606 pagesTreatment of Dry Skin SyndromePhuongNguyenNo ratings yet
- People in CleanroomsDocument13 pagesPeople in CleanroomsshdphNo ratings yet
- 2.1 The Integument Composed of Skin and Its Appendages, Sweat Glands, Sebaceous Glands, HairDocument6 pages2.1 The Integument Composed of Skin and Its Appendages, Sweat Glands, Sebaceous Glands, HairNuruel SazaNo ratings yet
- Multiple MyelomaDocument17 pagesMultiple MyelomaAhmed AbouelwafaNo ratings yet
- Anatomy of The Integumentary SystemDocument5 pagesAnatomy of The Integumentary SystemSmol PadernalNo ratings yet
- The Slim Book of Health Pearls: Man the Barricades - The Story of the Immune SystemFrom EverandThe Slim Book of Health Pearls: Man the Barricades - The Story of the Immune SystemNo ratings yet
- Integumentary SystemDocument17 pagesIntegumentary SystemNurani AtikasariNo ratings yet
- NIH Public Access: The Skin MicrobiomeDocument22 pagesNIH Public Access: The Skin MicrobiomeeaudreyliaNo ratings yet
- 1 s2.0 S0952791521001199 MainDocument7 pages1 s2.0 S0952791521001199 MainTatta CamposNo ratings yet
- Skin Microbiota: Majalah Kedokteran Sriwijaya, Th. 52 Nomor 1, January 2020Document6 pagesSkin Microbiota: Majalah Kedokteran Sriwijaya, Th. 52 Nomor 1, January 2020Rendra SyaniNo ratings yet
- Science Abo0693Document7 pagesScience Abo0693Lalo MunguiaNo ratings yet
- Biochemistryof SkinDocument17 pagesBiochemistryof Skinmigaloooo334No ratings yet
- Kamus ManusiaDocument822 pagesKamus ManusiaAhmad Abdullah100% (1)
- 2nd Half Microbiology For Allied Health StudentsDocument404 pages2nd Half Microbiology For Allied Health StudentsFjornNo ratings yet
- Anatomy, Skin (Integument) - StatPearls - NCBI BookshelfDocument4 pagesAnatomy, Skin (Integument) - StatPearls - NCBI Bookshelfputri aisheNo ratings yet
- Microbiota y DermatitisDocument6 pagesMicrobiota y Dermatitisdayenu barraNo ratings yet
- Sistema Inmune y PielDocument53 pagesSistema Inmune y PielKevin MendozaNo ratings yet
- HHS Public Access: Structure and Function of The Human Skin MicrobiomeDocument20 pagesHHS Public Access: Structure and Function of The Human Skin MicrobiomeEugeniaNo ratings yet
- Cetacean MorbilivirusDocument29 pagesCetacean MorbilivirusDio DvmNo ratings yet
- Mild AcidityDocument7 pagesMild AcidityDaniel Lee Eisenberg JacobsNo ratings yet
- Role of The Skin Microbiome in Dermatology: A LiteraturereviewDocument12 pagesRole of The Skin Microbiome in Dermatology: A LiteraturereviewIJAR JOURNALNo ratings yet
- XI Notes Immune System-2Document26 pagesXI Notes Immune System-2ramsha.wahid29No ratings yet
- Ijms 23 13071Document14 pagesIjms 23 13071indah permatasariNo ratings yet
- ANAPHY Integumentary SystemDocument3 pagesANAPHY Integumentary SystemNina JasminNo ratings yet
- Protective Mechanism of The Body (Mucous Membrane)Document3 pagesProtective Mechanism of The Body (Mucous Membrane)Kirk CobainNo ratings yet
- Stau Dinger 2011Document9 pagesStau Dinger 2011inforNo ratings yet
- Sarcoptes Scabiei: Mites Modulate Gene Expression in Human Skin EquivalentsDocument11 pagesSarcoptes Scabiei: Mites Modulate Gene Expression in Human Skin EquivalentsNailahRahmahNo ratings yet
- Toxicity of SkinDocument10 pagesToxicity of SkinMadhu Kiran DhondalayNo ratings yet
- Non-Specific Host Defenses Against Microbial PathogensDocument5 pagesNon-Specific Host Defenses Against Microbial PathogensSakil MunnaNo ratings yet
- The Keratinocyte As A Crucial Cell in The Predisposition, Onset, Progression, Therapy and Study of The Atopic DermatitisDocument30 pagesThe Keratinocyte As A Crucial Cell in The Predisposition, Onset, Progression, Therapy and Study of The Atopic Dermatitisvanessa_werbickyNo ratings yet
- TDDSDocument110 pagesTDDSp.ishaanpawarNo ratings yet
- Microbiome and Skin Biology: ReviewDocument6 pagesMicrobiome and Skin Biology: ReviewLinhNguyeNo ratings yet
- 33prozesky1982Document10 pages33prozesky1982Keto PrehranaNo ratings yet
- Human physiology: 3 main circuits and body's defense systemsDocument12 pagesHuman physiology: 3 main circuits and body's defense systemsYash BansodNo ratings yet
- Skin MicrobiotaDocument4 pagesSkin MicrobiotaDianaNo ratings yet
- Chapter 2Document26 pagesChapter 2cabdinuux32No ratings yet
- Structure of SkinDocument31 pagesStructure of Skinshaiknafisa1255No ratings yet
- Wnofns 43 (2022) 11-37Document27 pagesWnofns 43 (2022) 11-37Dennis StwnNo ratings yet
- The Skin MicrobiotaDocument11 pagesThe Skin MicrobiotaMélany Vélez ÁlavaNo ratings yet
- Characteristics of Living Organisms and The Integumentary SystemDocument11 pagesCharacteristics of Living Organisms and The Integumentary SystemSHYRE COLEEN CATAYAONo ratings yet
- Microbiom Skin VariabilityDocument5 pagesMicrobiom Skin VariabilityDianaNo ratings yet
- Burn Injury ICU CareDocument23 pagesBurn Injury ICU CareANGELICA MACASONo ratings yet
- Wound Bioburden: (To Fulfill The Task of Wound Care Subject)Document24 pagesWound Bioburden: (To Fulfill The Task of Wound Care Subject)Fransiska Marchelina HutabaratNo ratings yet
- Anatomy and Physiology of Sweat GlandDocument43 pagesAnatomy and Physiology of Sweat GlandUjjwala VermaNo ratings yet
- Limson - Human AnatomyDocument20 pagesLimson - Human AnatomyFaye RabulanNo ratings yet
- 2017 - Tecnología CosméticaDocument54 pages2017 - Tecnología CosméticaCésar Rosado Avila ToñoNo ratings yet
- Microorganisms 09 00543 v2Document19 pagesMicroorganisms 09 00543 v2Pande Agung MahariskiNo ratings yet
- Schwartz's Principles of Surgery: CH16: Page 1Document13 pagesSchwartz's Principles of Surgery: CH16: Page 1Kathryn ReunillaNo ratings yet
- The Role of Tight Junctions in Skin Barrier Function Anddermal AbsorptionDocument14 pagesThe Role of Tight Junctions in Skin Barrier Function Anddermal AbsorptionPaolaNo ratings yet
- Lec 1Document24 pagesLec 1Ab AbNo ratings yet
- The Integumentary SystemDocument21 pagesThe Integumentary SystemBella Agiussela0% (3)
- Normal Anatomy and Physiology-BurnsDocument4 pagesNormal Anatomy and Physiology-BurnsJusterine Jade To-ong SiglosNo ratings yet
- Biomolecules: Advanced Hydrogels As Wound DressingsDocument20 pagesBiomolecules: Advanced Hydrogels As Wound DressingsAndrian dwi yuliorofiNo ratings yet
- FungiDocument19 pagesFungimacalinabdiwali007No ratings yet
- Tobin Journal of Tissue ViabilityDocument18 pagesTobin Journal of Tissue ViabilityMonicaNo ratings yet
- 1 s2.0 S0022202X15447736 MainDocument7 pages1 s2.0 S0022202X15447736 MainKhan FatemaNo ratings yet
- Cosmeceuticals: Peptides, Proteins, and Growth Factors: Review ArticleDocument6 pagesCosmeceuticals: Peptides, Proteins, and Growth Factors: Review ArticleLeandro QuitoNo ratings yet
- Jaad 2021 Photoprotection-220823-165027Document21 pagesJaad 2021 Photoprotection-220823-165027Leandro QuitoNo ratings yet
- Impactoflightonskinhealth - 2022 220823 165027Document11 pagesImpactoflightonskinhealth - 2022 220823 165027Leandro QuitoNo ratings yet
- Neale2019 - SUNSCREENVITD 220823 165027Document9 pagesNeale2019 - SUNSCREENVITD 220823 165027Leandro QuitoNo ratings yet
- JCD - Nanosomesliposomes 220823 163443Document11 pagesJCD - Nanosomesliposomes 220823 163443Leandro QuitoNo ratings yet
- Oral Nail Growth Supplements: A Comprehensive ReviewDocument7 pagesOral Nail Growth Supplements: A Comprehensive ReviewLeandro QuitoNo ratings yet
- Hair Cosmetics For The Hair Loss Patient: Maria Fernanda Reis Gavazzoni Dias Aline Falci Loures Chloe EkelemDocument7 pagesHair Cosmetics For The Hair Loss Patient: Maria Fernanda Reis Gavazzoni Dias Aline Falci Loures Chloe EkelemLeandro QuitoNo ratings yet
- Novel Treatments - Research ArticleDocument7 pagesNovel Treatments - Research ArticleLeandro QuitoNo ratings yet
- El Rol Del Sensor de CalcioDocument10 pagesEl Rol Del Sensor de CalcioLeandro QuitoNo ratings yet
- Polymorphic Eruption of Pregnancy: ReviewDocument7 pagesPolymorphic Eruption of Pregnancy: ReviewLeandro QuitoNo ratings yet
- Desordenes Del Calcio AtpasaDocument7 pagesDesordenes Del Calcio AtpasaLeandro QuitoNo ratings yet
- Golgi Ca2+ Pump Activity Essential for Ca2+ Signaling and Viability in Darier Disease KeratinocytesDocument9 pagesGolgi Ca2+ Pump Activity Essential for Ca2+ Signaling and Viability in Darier Disease KeratinocytesLeandro QuitoNo ratings yet
- The Company of Biologists LTD J Cell Sci 2006 119:e401Document1 pageThe Company of Biologists LTD J Cell Sci 2006 119:e401Leandro QuitoNo ratings yet
- Scientific Program for Dermatology ConferenceDocument20 pagesScientific Program for Dermatology ConferenceLeandro QuitoNo ratings yet
- A Case-Control Study of Polymorphic Eruption of Pregnancy: A B C B ADocument5 pagesA Case-Control Study of Polymorphic Eruption of Pregnancy: A B C B ALeandro QuitoNo ratings yet
- This Holiday Was GreatDocument1 pageThis Holiday Was GreatLeandro QuitoNo ratings yet
- This Holiday Was GreatDocument1 pageThis Holiday Was GreatLeandro QuitoNo ratings yet
- Sanador - Lista Preturi Analize Medicale 2018Document24 pagesSanador - Lista Preturi Analize Medicale 2018AlinaIordacheNo ratings yet
- Our BerkshireTimes Magazine, Dec 2016 - Feb 2107Document24 pagesOur BerkshireTimes Magazine, Dec 2016 - Feb 2107Kathy Irene ReganNo ratings yet
- ContinueDocument2 pagesContinueShallu ManchandaNo ratings yet
- Gluten Determination From Foods: Figura 1. Classification of Prolamin SuperfamilyDocument6 pagesGluten Determination From Foods: Figura 1. Classification of Prolamin SuperfamilyanaNo ratings yet
- SP1ES EGRA Class Reading Profile Grade 2Document2 pagesSP1ES EGRA Class Reading Profile Grade 2Camille Langit DelacruzNo ratings yet
- Stress DisorderDocument33 pagesStress DisorderGwen PhamNo ratings yet
- (Pds Patklin) : Perhimpunan Dokter Spesialis Patologi Klinik Dan Kedokteran Laboratorium IndonesiaDocument5 pages(Pds Patklin) : Perhimpunan Dokter Spesialis Patologi Klinik Dan Kedokteran Laboratorium IndonesiaMariaNo ratings yet
- The Immune System: Warm UpDocument8 pagesThe Immune System: Warm UpvipNo ratings yet
- Ijhoscr: ABO Blood Grouping Mismatch in Hematopoietic Stem Cell Transplantation and Clinical GuidesDocument7 pagesIjhoscr: ABO Blood Grouping Mismatch in Hematopoietic Stem Cell Transplantation and Clinical GuidesMohana ReddyNo ratings yet
- Ha Ema To PoiesisDocument38 pagesHa Ema To PoiesisEmma Joel OtaiNo ratings yet
- Microvascular Research: SciencedirectDocument5 pagesMicrovascular Research: SciencedirectYunita AmiliaNo ratings yet
- Recognition of Immunodeficiency in The Newborn Period - UpToDateDocument21 pagesRecognition of Immunodeficiency in The Newborn Period - UpToDateAdela AlmadaNo ratings yet
- BCSC1718 S09 PDFDocument15 pagesBCSC1718 S09 PDFrisaNo ratings yet
- Vaccines: By: R.PH Dr. Saba Inayat Ali Lecturer Dow College of PharmacyDocument48 pagesVaccines: By: R.PH Dr. Saba Inayat Ali Lecturer Dow College of Pharmacyimran aliNo ratings yet
- Major Histocompatibility Complex (MHC) Structure and FunctionDocument16 pagesMajor Histocompatibility Complex (MHC) Structure and FunctionnikitaNo ratings yet
- Chapter 19 Blood 2304Document26 pagesChapter 19 Blood 2304Nishchaya SinghNo ratings yet
- CAP TODAY | MONOCYTE, IMMATUREDocument1 pageCAP TODAY | MONOCYTE, IMMATUREIlinca100% (1)
- Human Inborn Errors of Immunity: An Expanding Universe: ReviewDocument17 pagesHuman Inborn Errors of Immunity: An Expanding Universe: ReviewCony GSNo ratings yet
- Non-Specific Immune Response, and Resistance To Vibrio Alginolyticus in Pacific White, Chang 2012Document7 pagesNon-Specific Immune Response, and Resistance To Vibrio Alginolyticus in Pacific White, Chang 2012afandi saputraNo ratings yet
- Nure Seminar 0910114238Document38 pagesNure Seminar 0910114238ABAYNEGETAHUN getahunNo ratings yet
- Week 17 - THEORIES OF AGING - EACDocument44 pagesWeek 17 - THEORIES OF AGING - EACPaul Deliyos100% (1)
- A Comprehensive Review of Probiotics and Their Uses For Control of Viral Infections in The Wake of Pandemic Covid-19Document14 pagesA Comprehensive Review of Probiotics and Their Uses For Control of Viral Infections in The Wake of Pandemic Covid-19TJPLS Journal-Tropical Journal of Pharmaceutical and Life SciencesNo ratings yet