You might also like
- Weaver - ProbabilityDocument4 pagesWeaver - ProbabilityMauro SellittoNo ratings yet
- Probability Models for Pattern Recognition DecisionsDocument31 pagesProbability Models for Pattern Recognition Decisionsbobgalee100% (1)
- Module 5 RM Hypothesis TestingDocument21 pagesModule 5 RM Hypothesis TestingEm JayNo ratings yet
- The Theory of Evolution is a Result of Erroneous ExtrapolationFrom EverandThe Theory of Evolution is a Result of Erroneous ExtrapolationNo ratings yet
- Statistical Models ExplainedDocument5 pagesStatistical Models ExplainedBaba GhanushNo ratings yet
- Gambler's FallacyDocument11 pagesGambler's Fallacyprashantsahu20078549No ratings yet
- (Kahneman-Tversky) Belief in The Law of Small Numbers PDFDocument6 pages(Kahneman-Tversky) Belief in The Law of Small Numbers PDFaitor100% (1)
- Clinical Decision-Making: Learning ObjectivesDocument31 pagesClinical Decision-Making: Learning Objectivessavvy_as_98-1No ratings yet
- Hypothesis TestingDocument19 pagesHypothesis TestingtsrforfunNo ratings yet
- The Evolution of Diversity: How Evolution Generates Tremendous DiversityDocument14 pagesThe Evolution of Diversity: How Evolution Generates Tremendous DiversityAnkit JainNo ratings yet
- Probability in Bio-Sci 107-GENETICSDocument11 pagesProbability in Bio-Sci 107-GENETICSRico BelledoNo ratings yet
- Born This Way: Becoming, Being, and Understanding Scientists. Part 2: The Practice of Science and the Rise of Molecular BiologyFrom EverandBorn This Way: Becoming, Being, and Understanding Scientists. Part 2: The Practice of Science and the Rise of Molecular BiologyNo ratings yet
- HW 1Document3 pagesHW 1S SNo ratings yet
- Chapter 9 Biasses in Probability AssessmentDocument4 pagesChapter 9 Biasses in Probability AssessmentI Nyoman Sujana GiriNo ratings yet
- Biostatistics Module 7Document5 pagesBiostatistics Module 7JANE CASSANONo ratings yet
- Limits To Natural SelectionDocument10 pagesLimits To Natural SelectionKeren EspinozaNo ratings yet
- Evolutionary Route To Diploidy and SexDocument4 pagesEvolutionary Route To Diploidy and SexRogério Ribeiro MacêdoNo ratings yet
- Biostatistics Module 7 8Document10 pagesBiostatistics Module 7 8JANE CASSANONo ratings yet
- Advanced NDE Assignment 1 QuestionsDocument1 pageAdvanced NDE Assignment 1 QuestionsMathew JohnNo ratings yet
- Statistics 3 - ProbabilityDocument31 pagesStatistics 3 - Probabilityأمجد هاتف منفي جفالNo ratings yet
- Species: Understanding The Mathematics of DriftDocument8 pagesSpecies: Understanding The Mathematics of DriftTamara DrljačaNo ratings yet
- PropabilityDocument2 pagesPropabilityMohamed Omer MusaNo ratings yet
- The Originality of Species: Science and logic contradict the theory of evolution.From EverandThe Originality of Species: Science and logic contradict the theory of evolution.No ratings yet
- The Hot Hand Fallacy and The Gambler's Fallacy: Two Faces of Subjective Randomness?Document10 pagesThe Hot Hand Fallacy and The Gambler's Fallacy: Two Faces of Subjective Randomness?Edoardo Edmund Cohen CamassaNo ratings yet
- MMPC-5 ImpDocument32 pagesMMPC-5 ImpRajith KNo ratings yet
- Chapter 6: How To Avoid Accumulating Risk in Simple ComparisonsDocument9 pagesChapter 6: How To Avoid Accumulating Risk in Simple ComparisonspcorgoNo ratings yet
- Title: How Consciousness Emerges From Ions: Authors: Peilei Liu AffiliationsDocument6 pagesTitle: How Consciousness Emerges From Ions: Authors: Peilei Liu AffiliationsAlyssa HarperNo ratings yet
- Pnas 2007 Gerrish 6266 71Document6 pagesPnas 2007 Gerrish 6266 71api-211640838No ratings yet
- Add Math Project Work 2016Document12 pagesAdd Math Project Work 2016邓心悦33% (3)
- The Mermaid's Tale Ptolemaic Genetics Epicycles of LobbyingDocument10 pagesThe Mermaid's Tale Ptolemaic Genetics Epicycles of LobbyingzttNo ratings yet
- Dr. Gerald Crabtree - Our Fragile Intellect, Part IDocument3 pagesDr. Gerald Crabtree - Our Fragile Intellect, Part ILeakSourceInfoNo ratings yet
- Gerald Crabtree - Trends in Genetics. Our Fragile IntellectDocument6 pagesGerald Crabtree - Trends in Genetics. Our Fragile IntellectDemocracia real YA100% (1)
- Probability with Permutations: An Introduction To Probability And CombinationsFrom EverandProbability with Permutations: An Introduction To Probability And CombinationsNo ratings yet
- 315H Handout ChiSquareDocument3 pages315H Handout ChiSquareShyam PopatNo ratings yet
- Probability Theory: Experiments, Sample Spaces, Events and Equally Likely ProbabilitiesDocument3 pagesProbability Theory: Experiments, Sample Spaces, Events and Equally Likely Probabilitiesdeeptiv2004No ratings yet
- History of ProbabilityDocument3 pagesHistory of ProbabilityJeffery Ooi100% (1)
- Work SampleDocument24 pagesWork SampleTeen TalkNo ratings yet
- An Introduction, Some Recent Research, and A Dash of HistoryDocument3 pagesAn Introduction, Some Recent Research, and A Dash of HistoryMitchel RodwellNo ratings yet
- Cloning and Embryonic Stem Cells: A New Era in Human Biology and MedicineDocument10 pagesCloning and Embryonic Stem Cells: A New Era in Human Biology and Medicine7tuihjkhNo ratings yet
- Understanding Cancer from a Systems Biology Point of View: From Observation to Theory and BackFrom EverandUnderstanding Cancer from a Systems Biology Point of View: From Observation to Theory and BackRating: 5 out of 5 stars5/5 (4)
- What Is Random?: Chance and Order in Mathematics and LifeFrom EverandWhat Is Random?: Chance and Order in Mathematics and LifeRating: 3.5 out of 5 stars3.5/5 (2)
- Research Paper Outline On Human CloningDocument8 pagesResearch Paper Outline On Human Cloninggz98szx1No ratings yet
- Problem Set 3Document3 pagesProblem Set 3Kylie KimNo ratings yet
- MosaicsmDocument8 pagesMosaicsmLucas TobingNo ratings yet
- Approaches To ProbabilityDocument4 pagesApproaches To ProbabilitybhishmNo ratings yet
- Xienadin Group Project - Final OutputDocument30 pagesXienadin Group Project - Final OutputReshiel NicolasNo ratings yet
- DNA Mutation RatesDocument37 pagesDNA Mutation RatesSean Pitman, MDNo ratings yet
- Session 1 Basic Concepts and Principles in Statistics and Probability-Part 2Document31 pagesSession 1 Basic Concepts and Principles in Statistics and Probability-Part 2EDDIE T. PEREZNo ratings yet
- Handout 1-Foundations of Probability TheoryDocument27 pagesHandout 1-Foundations of Probability Theorylwandle221No ratings yet
- Capítulo 3 - RosnerDocument13 pagesCapítulo 3 - RosnerJasiel CabreraNo ratings yet
- IELTS PracticeDocument6 pagesIELTS PracticeSabella XenaNo ratings yet
- Intro To Stat (STAT 111) by EwensDocument113 pagesIntro To Stat (STAT 111) by EwensIris ZhangNo ratings yet
- Population Genetics and The HardyDocument5 pagesPopulation Genetics and The HardyjelkicalelaNo ratings yet
- Exercises Sheet 1 Types of Statistical Data and Display of DataDocument10 pagesExercises Sheet 1 Types of Statistical Data and Display of DataJaime-Nemilyn Ati-id FadcharNo ratings yet
- Inductive Reasoning and Chance Discovery : Ó 2004 Kluwer Academic Publishers. Printed in The NetherlandsDocument11 pagesInductive Reasoning and Chance Discovery : Ó 2004 Kluwer Academic Publishers. Printed in The NetherlandsVíctor Eduardo Hernández BenavidesNo ratings yet
- Math - Subjuctive Probability MeaningDocument3 pagesMath - Subjuctive Probability MeaningAloysius RajNo ratings yet
- Genetics Brain Teasers: Explore your Understadning of Genes, DNA, RNA, mRNA, Chromosomes, Mutation, Heredity, Evolution, and more!From EverandGenetics Brain Teasers: Explore your Understadning of Genes, DNA, RNA, mRNA, Chromosomes, Mutation, Heredity, Evolution, and more!No ratings yet
- BioEssays Volume 9 Issue 2-3 1988 (Doi 10.1002/bies.950090212) Robert A. Weinberg - The Human Genome Sequence - What Will It Do For Us PDFDocument2 pagesBioEssays Volume 9 Issue 2-3 1988 (Doi 10.1002/bies.950090212) Robert A. Weinberg - The Human Genome Sequence - What Will It Do For Us PDFnadia_676766No ratings yet
- Gene MappingDocument14 pagesGene MappingPROFESSORNo ratings yet
- Life Science Cell Theory and MicroscopyDocument16 pagesLife Science Cell Theory and MicroscopyElhade ElbakushNo ratings yet
- DNA Kinetics ReassociationDocument4 pagesDNA Kinetics ReassociationSarmistha NayakNo ratings yet
- Modul AdjectiveDocument5 pagesModul AdjectiveFadli Nora IrandaNo ratings yet
- Lesson Plan GuideDocument5 pagesLesson Plan Guideapi-665220631No ratings yet
- Sexual Reproduction in Flowering PlantsDocument4 pagesSexual Reproduction in Flowering PlantsHarpreetNo ratings yet
- 10 1089@omi 2019 0192Document12 pages10 1089@omi 2019 0192Raul Morales VillegasNo ratings yet
- PMLS Lesson 5Document7 pagesPMLS Lesson 5Althea EspirituNo ratings yet
- AP Biology Reading Guide Chapter 23Document12 pagesAP Biology Reading Guide Chapter 23shkjsdhfskjd60% (5)
- Issue On Human DevelopmentDocument15 pagesIssue On Human DevelopmentJhaga PotpotNo ratings yet
- Evidence for Evolution of WhalesDocument6 pagesEvidence for Evolution of WhalesMya IrelandNo ratings yet
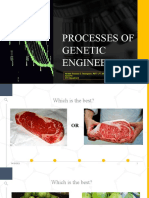
- Processes of Genetic Engineering: Noraine Princess G. Tabangcora, RMT, LPT, MSMT Asnshs SHS DepartmentDocument25 pagesProcesses of Genetic Engineering: Noraine Princess G. Tabangcora, RMT, LPT, MSMT Asnshs SHS DepartmentKayla Mae GaNo ratings yet
- Instruction Dignitas Personae On Certain Bioethical Questions, Congregation For The Doctrine of The FaithDocument19 pagesInstruction Dignitas Personae On Certain Bioethical Questions, Congregation For The Doctrine of The FaithJBSfanNo ratings yet
- Cambridge International General Certificate of Secondary EducationDocument16 pagesCambridge International General Certificate of Secondary EducationZaigham ZiaNo ratings yet
- Patterns of Inheritance Multiple Choice QuizDocument16 pagesPatterns of Inheritance Multiple Choice QuizjevaireNo ratings yet
- RK PandeyDocument211 pagesRK PandeyNirdeshKumarSharmaNo ratings yet
- Pikachurin: EGFLAM GeneDocument6 pagesPikachurin: EGFLAM Genefelipe smithNo ratings yet
- Lesson Plan in Cells (Demo)Document3 pagesLesson Plan in Cells (Demo)Gerald Galvez ApostolNo ratings yet
- Microarray Technology - A Brief Introduction - : Markus PanhuysenDocument38 pagesMicroarray Technology - A Brief Introduction - : Markus PanhuysenrashmiNo ratings yet
- Unit 1: Historical SourcesDocument33 pagesUnit 1: Historical SourcesMUSIRIKARENo ratings yet
- Reviewer (Gen. Biology 2)Document5 pagesReviewer (Gen. Biology 2)ianah aloriaNo ratings yet
- WEEK 3 LAB EXERCISE - Cell Structures and Functions - UY-OCODocument4 pagesWEEK 3 LAB EXERCISE - Cell Structures and Functions - UY-OCOBianca LouiseNo ratings yet
- Earl Eugene Castro - Bio 32 Learning Activity 1Document2 pagesEarl Eugene Castro - Bio 32 Learning Activity 1EARL CASTRONo ratings yet
- The Mixed-Up Brothers of Bogotá - The New York TimesDocument33 pagesThe Mixed-Up Brothers of Bogotá - The New York TimesLakeisha Christi LalasNo ratings yet
- Cell TheoryDocument2 pagesCell TheoryAnya SharmaNo ratings yet
- Olivera 2002Document29 pagesOlivera 2002Nathali GalindoNo ratings yet
- General Problem Solving With Search Algorithms: 11/24/20 (C) Debasis Mitra 1Document52 pagesGeneral Problem Solving With Search Algorithms: 11/24/20 (C) Debasis Mitra 1Sheelaj BabuNo ratings yet
- 1 2021 Plant Tissue Culture in Horticultural CropsDocument4 pages1 2021 Plant Tissue Culture in Horticultural CropsCarlos Campuzano CalvoNo ratings yet
- Environmental Biotechnology - BBTechDocument5 pagesEnvironmental Biotechnology - BBTechBekele OljiraNo ratings yet
- 6th Science PDE Worksheet 2018 1 TermDocument5 pages6th Science PDE Worksheet 2018 1 TermJUAN CARLOS0% (1)