You might also like
- Nutrition and Enhanced Sports Performance: Muscle Building, Endurance, and StrengthFrom EverandNutrition and Enhanced Sports Performance: Muscle Building, Endurance, and StrengthRating: 4 out of 5 stars4/5 (4)
- (2015) Exercise and SarcopeniaDocument3 pages(2015) Exercise and SarcopeniaTatiane vitorNo ratings yet
- Nutrition and Skeletal MuscleFrom EverandNutrition and Skeletal MuscleStéphane WalrandNo ratings yet
- Leucine Regulates Translation Initiation of Protein Synthesis in Skeletal Muscle After ExerciseDocument6 pagesLeucine Regulates Translation Initiation of Protein Synthesis in Skeletal Muscle After ExerciseLaurensius45 NugrohoNo ratings yet
- Asam Amino LeusinDocument5 pagesAsam Amino LeusinIceBearNo ratings yet
- 6 - Dietary Protein For Muscle HypertrophyDocument2 pages6 - Dietary Protein For Muscle HypertrophyJéssica LagoNo ratings yet
- 6 - Dietary Protein For Muscle HypertrophyDocument2 pages6 - Dietary Protein For Muscle HypertrophyJéssica LagoNo ratings yet
- Protein Requirements and Supplementation in Strength Sports: Nutrition July 2004Document8 pagesProtein Requirements and Supplementation in Strength Sports: Nutrition July 2004Andrei ChirilaNo ratings yet
- Dietary Protein, Endurance Exercise, and Human Skeletal-Muscle Protein TurnoverDocument6 pagesDietary Protein, Endurance Exercise, and Human Skeletal-Muscle Protein TurnoverRosemberg PriscoNo ratings yet
- Endocrine Responses and Acute mTOR Pathway Phosphorylation To Resistance Exercise With Leucine and WheyDocument7 pagesEndocrine Responses and Acute mTOR Pathway Phosphorylation To Resistance Exercise With Leucine and WheyIsa AguilarNo ratings yet
- Symposium 2: Exercise and Protein Nutrition The Science of Muscle Hypertrophy: Making Dietary Protein CountDocument4 pagesSymposium 2: Exercise and Protein Nutrition The Science of Muscle Hypertrophy: Making Dietary Protein Countivan pitrulliNo ratings yet
- Nxy 263Document10 pagesNxy 263Conciencia CristalinaNo ratings yet
- Per I ExerciseDocument8 pagesPer I ExerciseJesus Rafael Zapata AlvarezNo ratings yet
- Sintese Proteica e Fontes de ProteinaDocument11 pagesSintese Proteica e Fontes de ProteinanutrinathyalvesNo ratings yet
- Symposium 2: Exercise and Protein Nutrition The Science of Muscle Hypertrophy: Making Dietary Protein CountDocument4 pagesSymposium 2: Exercise and Protein Nutrition The Science of Muscle Hypertrophy: Making Dietary Protein CountNoJsterNo ratings yet
- mTOR Signaling Pathway and Myostatin in Men Resistance Exercise With Whey Protein Ingestion AffectsDocument11 pagesmTOR Signaling Pathway and Myostatin in Men Resistance Exercise With Whey Protein Ingestion AffectsMarcosViníciusSilvaNo ratings yet
- Activation of mTOR Signalling in Young and Old Human Skeletal Muscle in Response To Combined Resistance Exercise and Whey Protein IngestionDocument10 pagesActivation of mTOR Signalling in Young and Old Human Skeletal Muscle in Response To Combined Resistance Exercise and Whey Protein IngestionCarlos RodriguezNo ratings yet
- Ramping Up The Signal: Promoting Endurance Training Adaptation in Skeletal Muscle by Nutritional ManipulationDocument7 pagesRamping Up The Signal: Promoting Endurance Training Adaptation in Skeletal Muscle by Nutritional ManipulationFernando IsaacNo ratings yet
- Nutrition For SprinterDocument18 pagesNutrition For SprinterscribdeseNo ratings yet
- Nutritional Interventions To Promote PosDocument191 pagesNutritional Interventions To Promote PosFelipe FerreiraNo ratings yet
- Effects of Methoxyisoflavone, Ecdysterone, and Sulfo-Polysaccharide Supplementation On Training Adaptations in Resistance-Trained MalesDocument9 pagesEffects of Methoxyisoflavone, Ecdysterone, and Sulfo-Polysaccharide Supplementation On Training Adaptations in Resistance-Trained Malespuran1234567890No ratings yet
- Protein Requirements For Athletes: TrainingDocument2 pagesProtein Requirements For Athletes: Trainingapi-58002895No ratings yet
- Effects of Protein Intake Prior To Carbohydrate-Restricted Endurance Exercise A Randomized Crossover TrialDocument13 pagesEffects of Protein Intake Prior To Carbohydrate-Restricted Endurance Exercise A Randomized Crossover TrialJuan Luis RuizNo ratings yet
- Nutrition For The SprinterDocument12 pagesNutrition For The Sprinterhrvoje09No ratings yet
- PROGENEX Recovery Tech ArticleDocument5 pagesPROGENEX Recovery Tech ArticlePROGENEXNo ratings yet
- Close+etal ,+2016+New+StrategiasDocument3 pagesClose+etal ,+2016+New+StrategiasVanessa BarrosNo ratings yet
- New Strategies in Sport NutritionDocument15 pagesNew Strategies in Sport NutritionagcNo ratings yet
- Kraemer 2009 HMB 00005768-200905000-00018Document11 pagesKraemer 2009 HMB 00005768-200905000-00018ewlfitnessconsultingNo ratings yet
- Is There A Need For Protein Ingestion During Exercise?Document7 pagesIs There A Need For Protein Ingestion During Exercise?aguaschma2114No ratings yet
- Branched Chain Amino Acids and Muscle Protein Synthesis in Humans Myth or RealityDocument8 pagesBranched Chain Amino Acids and Muscle Protein Synthesis in Humans Myth or RealityY Tran Trang MyNo ratings yet
- Independent and Combined Effects of Amino Acids and Glucose After Resistance ExerciseDocument7 pagesIndependent and Combined Effects of Amino Acids and Glucose After Resistance ExerciseIgor FernandesNo ratings yet
- Amilopectina + Whey Vip 2017Document9 pagesAmilopectina + Whey Vip 2017Diana PrietoNo ratings yet
- Nutrition For The SprinterDocument12 pagesNutrition For The SprinterMarcelo NogueraNo ratings yet
- Co-Ingestion of Nutritional Ergogenic Aids and High-IntensityDocument12 pagesCo-Ingestion of Nutritional Ergogenic Aids and High-IntensityVanessa BarrosNo ratings yet
- Nutrients: Effects of Beta-Alanine On Muscle Carnosine and Exercise Performance: A Review of The Current LiteratureDocument23 pagesNutrients: Effects of Beta-Alanine On Muscle Carnosine and Exercise Performance: A Review of The Current Literatureseggy7No ratings yet
- Low-Carbohydrate Training Increases Protein Requirements of Endurance AthletesDocument8 pagesLow-Carbohydrate Training Increases Protein Requirements of Endurance AthletesAlejandro Estrada RiosNo ratings yet
- Protein QualityDocument9 pagesProtein QualityAvanti TorNo ratings yet
- Protein Blend Ingestion Following Resistance Exercise Promotes Human Muscle Protein SynthesisDocument7 pagesProtein Blend Ingestion Following Resistance Exercise Promotes Human Muscle Protein SynthesisAnderson Anaya TreinadorNo ratings yet
- 1550 2783 3 1 7 PDFDocument21 pages1550 2783 3 1 7 PDFlucasNo ratings yet
- Protein Hydrolysates, Myogenia 2016 V1.0Document11 pagesProtein Hydrolysates, Myogenia 2016 V1.0Stefanie MacherhammerNo ratings yet
- And Christopher J. Lynch Pengxiang She, Yingsheng Zhou, Zhiyou Zhang, Kathleen Griffin, Kavitha GowdaDocument10 pagesAnd Christopher J. Lynch Pengxiang She, Yingsheng Zhou, Zhiyou Zhang, Kathleen Griffin, Kavitha Gowdaglico10No ratings yet
- Effects of Methoxyisoflavone, Ecdysterone, and Sulfo-Polysaccharide Supplementation On Training Adaptations in Resistance-Trained MalesDocument9 pagesEffects of Methoxyisoflavone, Ecdysterone, and Sulfo-Polysaccharide Supplementation On Training Adaptations in Resistance-Trained Malesaitor garciaNo ratings yet
- Gonzalez 2015Document15 pagesGonzalez 2015Mário SimõesNo ratings yet
- knh304 Research PaperDocument11 pagesknh304 Research Paperapi-384481487No ratings yet
- 2009 - Rational For Protein Blen in Sports NutritionDocument9 pages2009 - Rational For Protein Blen in Sports NutritionAllan ScottNo ratings yet
- LAT1 Protein Content Increases Following 12 WeeksDocument15 pagesLAT1 Protein Content Increases Following 12 WeeksRosaneLacerdaNo ratings yet
- Jurnal InterDocument11 pagesJurnal InterEva IndrianiNo ratings yet
- Human Skeletal Muscle Protein Metabolism Responses To Amino Acid NutritionDocument11 pagesHuman Skeletal Muscle Protein Metabolism Responses To Amino Acid NutritionNisaNo ratings yet
- The Role of Milk - and Soy-Based Protein in Support of Muscle Protein Synthesis and Muscle Protein Accretion in Young and Elderly PersonsDocument12 pagesThe Role of Milk - and Soy-Based Protein in Support of Muscle Protein Synthesis and Muscle Protein Accretion in Young and Elderly Personsscribder100% (1)
- JSCR 20 3 643 653 06Document12 pagesJSCR 20 3 643 653 06Deepak SinghNo ratings yet
- The Relation Between Exercise Duration and Intensity PCRDocument4 pagesThe Relation Between Exercise Duration and Intensity PCRKlaraNo ratings yet
- 10 1152@japplphysiol 00907 2010Document11 pages10 1152@japplphysiol 00907 2010saulo freitas pereiraNo ratings yet
- BETA ALANINA the Effect of Β-Alanine Supplementation During High-Intensity Interval Training on 3 Repeated Sprint Ability Performance and Neuromuscular FatigueDocument40 pagesBETA ALANINA the Effect of Β-Alanine Supplementation During High-Intensity Interval Training on 3 Repeated Sprint Ability Performance and Neuromuscular Fatiguejanailma gomesNo ratings yet
- Nutrients 12 02023 v2Document4 pagesNutrients 12 02023 v2markis grcNo ratings yet
- The Effects of Nutritional SupplementatiDocument12 pagesThe Effects of Nutritional SupplementatiFelipe FerreiraNo ratings yet
- Atualização em Nutrição EsportivaDocument42 pagesAtualização em Nutrição EsportivaHumberto GomesNo ratings yet
- Schoenfeld 2013Document16 pagesSchoenfeld 2013Jean Freitas LimaNo ratings yet
- Mechanisms of Metabolic Stress in Hypertrophi - Brad J. SchoenfeldDocument18 pagesMechanisms of Metabolic Stress in Hypertrophi - Brad J. SchoenfeldCesar Martin Alonso100% (1)
- Effects of Protein Supplements On Muscle Damage, Soreness and Recovery of Muscle Function and Physical..Document17 pagesEffects of Protein Supplements On Muscle Damage, Soreness and Recovery of Muscle Function and Physical..Amir HayatNo ratings yet
- Burke 2018Document8 pagesBurke 2018GerasimarasNo ratings yet
- Banana RND in MalaysiaDocument5 pagesBanana RND in MalaysiaChee'dida FelidaeNo ratings yet
- mcb130 Sci PaperDocument9 pagesmcb130 Sci PaperShannen SenaNo ratings yet
- Acute-Phase ProteinDocument17 pagesAcute-Phase ProteinAbhijith S. PNo ratings yet
- 2007 A Level H2 Biology P2 AnsDocument6 pages2007 A Level H2 Biology P2 AnsjoannetzyNo ratings yet
- NGM Detect User GuideDocument144 pagesNGM Detect User GuideFe LipeNo ratings yet
- Welch Et AlDocument2 pagesWelch Et AlStephanie SmithNo ratings yet
- Cyclins and Cyclin-Dependent Kinases (CDKS)Document16 pagesCyclins and Cyclin-Dependent Kinases (CDKS)Sudarshan KiniNo ratings yet
- Vietnam National University Ho Chi Minh International UniversityDocument4 pagesVietnam National University Ho Chi Minh International UniversityHuỳnh NhưNo ratings yet
- Chapter 1 Introduction To BiophysicsDocument17 pagesChapter 1 Introduction To BiophysicsNurarief Affendy100% (2)
- ELISA AssayDocument13 pagesELISA AssaynawazashNo ratings yet
- LAS Physical Science Week 4Document10 pagesLAS Physical Science Week 4Shekaina Faith Cuizon LozadaNo ratings yet
- Pathophysiology of Graft Versus Host DiseaseDocument2 pagesPathophysiology of Graft Versus Host DiseasePriMaNo ratings yet
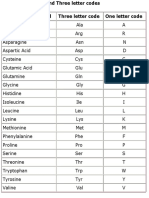
- Amino Acid CodesDocument1 pageAmino Acid Codesmorzan ortizNo ratings yet
- DNA TechnologyDocument27 pagesDNA TechnologyIM CTNo ratings yet
- What's in Breast Milk?Document1 pageWhat's in Breast Milk?Sunny SideNo ratings yet
- Subhashree Parida 2101020204-AssignmentDocument8 pagesSubhashree Parida 2101020204-AssignmentHero PratikNo ratings yet
- IL1 AntagonistaDocument13 pagesIL1 AntagonistaZitlal-lin VictoriaNo ratings yet
- Enzyme-Mediated DNA Looping - Halford, Welsh, Szczelkun - 2004Document26 pagesEnzyme-Mediated DNA Looping - Halford, Welsh, Szczelkun - 2004innu88No ratings yet
- Oxidative Stress and Antioxidant DefenseDocument558 pagesOxidative Stress and Antioxidant DefenseCLAUDIANo ratings yet
- BPS2110 Midterm 1 AnswersDocument5 pagesBPS2110 Midterm 1 AnswersMaya AkkawiNo ratings yet
- Bio 462 - Experiment 4Document3 pagesBio 462 - Experiment 4using formeNo ratings yet
- CHEM 151 (Chapter 4)Document4 pagesCHEM 151 (Chapter 4)Chantel Acevero100% (1)
- Amino Acid: Navigation SearchDocument11 pagesAmino Acid: Navigation SearchSamiuddin YezdaniNo ratings yet
- ViaguardDocument2 pagesViaguardviaguardNo ratings yet
- Physical Science M4Document20 pagesPhysical Science M4Aeron GallardoNo ratings yet
- Ribosomes and Protein SynthesisDocument15 pagesRibosomes and Protein SynthesisRichard ShakespeareNo ratings yet
- Dna PolymeraseDocument25 pagesDna PolymerasePoojaNo ratings yet
- Emboss PDFDocument54 pagesEmboss PDFFrancisco JavierNo ratings yet
- Dna and Rna Grade 10 TopicsDocument16 pagesDna and Rna Grade 10 TopicsJewel Tan100% (1)
- DR P Gupta ImtechDocument4 pagesDR P Gupta ImtechaaimpossibleNo ratings yet
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (516)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4 out of 5 stars4/5 (5)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (4)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineFrom EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineRating: 4 out of 5 stars4/5 (17)