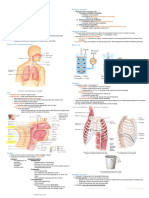
You might also like
- Sodr2017 Nova Especie CrotonDocument9 pagesSodr2017 Nova Especie CrotonShaline SefaraNo ratings yet
- DichorisandrastriatulaDocument9 pagesDichorisandrastriatulaProyecto Orquídeas BPAMNo ratings yet
- Luz Natural 2017Document11 pagesLuz Natural 2017Roger VenturaNo ratings yet
- Chemistry Biodiversity - 2023 - Galvão - Seasonal and Circadian Evaluation of The Essential Oil of Dizygostemon RipariusDocument9 pagesChemistry Biodiversity - 2023 - Galvão - Seasonal and Circadian Evaluation of The Essential Oil of Dizygostemon Ripariusan_marNo ratings yet
- 10 - 2017 - Melo Júnior Et Al. 2017 - Ciência & NaturaDocument10 pages10 - 2017 - Melo Júnior Et Al. 2017 - Ciência & NaturaÍgor Abba ArriolaNo ratings yet
- Ometric Morphometrics of Leaves of Anacardium Microcarpum Dan AoDocument13 pagesOmetric Morphometrics of Leaves of Anacardium Microcarpum Dan AofeNo ratings yet
- Plecevo Article 95601 en 1Document18 pagesPlecevo Article 95601 en 1Maurício GonçalvesNo ratings yet
- Ant Species (Hymenoptera, Formicidae) From The Seasonally Dry Tropical Forest of Northeastern Brazil: A Compilation From Field Surveys in Bahia and Literature RecordsDocument8 pagesAnt Species (Hymenoptera, Formicidae) From The Seasonally Dry Tropical Forest of Northeastern Brazil: A Compilation From Field Surveys in Bahia and Literature RecordsKean DeanNo ratings yet
- MICROALGAE STUDYDocument17 pagesMICROALGAE STUDYRenato JucianoNo ratings yet
- New Records of Utricularia (Lentibulariaceae) For The State of Maranhão, BrazilDocument6 pagesNew Records of Utricularia (Lentibulariaceae) For The State of Maranhão, BrazilMilena SalazarNo ratings yet
- Leaf Morphological Strategies of Seedlings and Saplings of Rhizophora Mangle (Rhizophoraceae), Laguncularia Racemosa (Combretaceae) andDocument14 pagesLeaf Morphological Strategies of Seedlings and Saplings of Rhizophora Mangle (Rhizophoraceae), Laguncularia Racemosa (Combretaceae) andAndressa PelozoNo ratings yet
- Rando Et Al. 2016Document16 pagesRando Et Al. 2016Isabel PinheiroNo ratings yet
- A Synopsis of The Rubiaceae of The States of Mato Grosso and Mato Grosso Do Sul, Brazil, With A Key To Genera, and A Preliminary Species ListDocument84 pagesA Synopsis of The Rubiaceae of The States of Mato Grosso and Mato Grosso Do Sul, Brazil, With A Key To Genera, and A Preliminary Species ListJúnior SenseyNo ratings yet
- Mastozoología Neotropical 0327-9383: Issn: Ulyses@cenpat - Edu.arDocument13 pagesMastozoología Neotropical 0327-9383: Issn: Ulyses@cenpat - Edu.arpaco jonesNo ratings yet
- Vegetative Characterization and Key of Myrtaceae Species From A Remnant of Araucaria Rainforest, Curitiba, ParanáDocument28 pagesVegetative Characterization and Key of Myrtaceae Species From A Remnant of Araucaria Rainforest, Curitiba, ParanáMarcus Vinicius Duarte RodriguesNo ratings yet
- 2 Macro PDFDocument22 pages2 Macro PDFLINA MARIA RODRIGUEZ RUIZNo ratings yet
- New Records of Macrofungi (Basidiomycota) For Colombia From A Tropical Dry ForestDocument22 pagesNew Records of Macrofungi (Basidiomycota) For Colombia From A Tropical Dry ForestLINA MARIA RODRIGUEZ RUIZNo ratings yet
- Brugmansia candida Extract Deters Hypsipyla grandellaDocument12 pagesBrugmansia candida Extract Deters Hypsipyla grandellaPaulkik22No ratings yet
- Brugmansia candida Extract Deters Hypsipyla grandellaDocument12 pagesBrugmansia candida Extract Deters Hypsipyla grandellaPaulkik22No ratings yet
- Ryegrass genotypes biomass and quality in southern BrazilDocument9 pagesRyegrass genotypes biomass and quality in southern BraziladrianoNo ratings yet
- Assessment of Coconut Tree Genetic Divergence by Compound Sample RAPD Marker AnalysisDocument8 pagesAssessment of Coconut Tree Genetic Divergence by Compound Sample RAPD Marker Analysisspullc26No ratings yet
- Leaf Area Estimation of Anacardium HumileDocument8 pagesLeaf Area Estimation of Anacardium HumileAngelita SoaresNo ratings yet
- 1 SM PDFDocument8 pages1 SM PDFLucas Meirelles SiqueiraNo ratings yet
- First Cytomolecular Characterization of Three Neot PDFDocument7 pagesFirst Cytomolecular Characterization of Three Neot PDFGABRIELA MONTEIRONo ratings yet
- CheckList Article 25463 en 1Document13 pagesCheckList Article 25463 en 1Francisco JoséNo ratings yet
- Rubiaceae Similarity and Conservation in Brazilian ForestsDocument13 pagesRubiaceae Similarity and Conservation in Brazilian ForestsJamesNo ratings yet
- Artropodos Del DoselDocument10 pagesArtropodos Del DoselMauricio GarciaNo ratings yet
- Mendes Et Al. 2017 - Reproductive Phenology of Mauritia Flexuosa L. (Arecaceae)Document9 pagesMendes Et Al. 2017 - Reproductive Phenology of Mauritia Flexuosa L. (Arecaceae)Yakov Mario QuinterosNo ratings yet
- 1 Martins - 2017 - Butterflies Amazon and Cerrado BrazilDocument11 pages1 Martins - 2017 - Butterflies Amazon and Cerrado BrazilIdes Daniela Herazo FdezNo ratings yet
- Injerto Tomate en Solanaceas - 2Document10 pagesInjerto Tomate en Solanaceas - 2Jeisson Daniel Torres QuinteroNo ratings yet
- Med Plants Brazil ForestDocument23 pagesMed Plants Brazil ForestChloe Devon Littell-SimpsonNo ratings yet
- Artigo LajedicolaDocument9 pagesArtigo LajedicolaPedro CardosoNo ratings yet
- 2004_Schäffer_Environmental_and_Ontogenetic_Control_ofDocument14 pages2004_Schäffer_Environmental_and_Ontogenetic_Control_ofmariangelestmNo ratings yet
- Artigo 1 InglesDocument10 pagesArtigo 1 InglesThiago LebraNo ratings yet
- Green Lacewings Associated with Melon Crop in BrazilDocument2 pagesGreen Lacewings Associated with Melon Crop in Brazilbolosphex1634No ratings yet
- Histopathology of Endocrine Organs of Miranda S White Lippe - 2022 - Journal ofDocument10 pagesHistopathology of Endocrine Organs of Miranda S White Lippe - 2022 - Journal ofmatheuscalixto.s999No ratings yet
- Acta Scientiarum 1Document11 pagesActa Scientiarum 1Manelli77No ratings yet
- Motta Tavaresetal.2016 EcologyofPhyllodytesluteolusDocument12 pagesMotta Tavaresetal.2016 EcologyofPhyllodytesluteolusvanessa e bruno becaciciNo ratings yet
- 0001 3765 Aabc 201520140340 PDFDocument8 pages0001 3765 Aabc 201520140340 PDFÁlvaro CarrasquelNo ratings yet
- Jorge Et Al 2020 - Herpetofauna Associada À Macambira-De-FlechaDocument12 pagesJorge Et Al 2020 - Herpetofauna Associada À Macambira-De-FlechaRaul SalesNo ratings yet
- 11.1.128-Grosso et al., 2022Document24 pages11.1.128-Grosso et al., 2022Diego BaldoNo ratings yet
- 01 Valões Araújo+Et+AlDocument11 pages01 Valões Araújo+Et+Alaedsp2No ratings yet
- Entomofauna Associated To Different Phenological Stages On Blueberry CropDocument13 pagesEntomofauna Associated To Different Phenological Stages On Blueberry CropSarai Olmedo CruzNo ratings yet
- 07 - ( - ARTIGO - ) - LIMA Et Al (2019) - New Records of Phallales (Basidiomycota) From Brazilian Semi-Arid RegionDocument10 pages07 - ( - ARTIGO - ) - LIMA Et Al (2019) - New Records of Phallales (Basidiomycota) From Brazilian Semi-Arid RegionRenato JucianoNo ratings yet
- Bromelicolous Habit of Three Species of Rhinella Fitzinger, 1826 (Anura: Bufonidae) in Northeastern Brazil's Atlantic ForestDocument4 pagesBromelicolous Habit of Three Species of Rhinella Fitzinger, 1826 (Anura: Bufonidae) in Northeastern Brazil's Atlantic Forestvanessa e bruno becaciciNo ratings yet
- Mendes-Silva Et Al. 2021 - Flora of Rondônia, Brazil Malmeeae (Annonaceae)Document27 pagesMendes-Silva Et Al. 2021 - Flora of Rondônia, Brazil Malmeeae (Annonaceae)Márcio BazanteNo ratings yet
- Journal of EthnopharmacologyDocument34 pagesJournal of EthnopharmacologygabrielledireitoNo ratings yet
- CHO in Leaves of Brazlian Grasses Moraes - Et - Al-2013-Grass - and - Forage - ScienceDocument13 pagesCHO in Leaves of Brazlian Grasses Moraes - Et - Al-2013-Grass - and - Forage - ScienceKalanNo ratings yet
- Ajabssp 2011 462 468Document7 pagesAjabssp 2011 462 468Julio ZacNo ratings yet
- Art.2 ExtraccionDocument13 pagesArt.2 ExtraccionAngela GuidoNo ratings yet
- Prosopanche CocucciiDocument16 pagesProsopanche CocucciiYasmin Vitória kastenNo ratings yet
- Cano Et Al - 2013 - A Revision of Trithrinax PDFDocument53 pagesCano Et Al - 2013 - A Revision of Trithrinax PDFHenoh100% (3)
- PharmacognJ 10 5 823 PDFDocument4 pagesPharmacognJ 10 5 823 PDFMr SetiawanNo ratings yet
- Kal An Choe Brasiliensis Cam BDocument19 pagesKal An Choe Brasiliensis Cam BReema RoyNo ratings yet
- Artigo ZoologiaDocument8 pagesArtigo ZoologiaCris SantosNo ratings yet
- Alternative Reproductive Modes of Atlantic Forest FrogsDocument6 pagesAlternative Reproductive Modes of Atlantic Forest FrogsWenner BritoNo ratings yet
- DownloadDocument10 pagesDownloadKatrin Choisy D. PascualNo ratings yet
- Maturação GuanandiDocument9 pagesMaturação GuanandiEvanny ValleriaNo ratings yet
- Isotopic Discrimination Factors (Δ 13C and Δ 15N) between Tissues and Diet of the Broad-Snouted Caiman (Caiman latirostris)Document7 pagesIsotopic Discrimination Factors (Δ 13C and Δ 15N) between Tissues and Diet of the Broad-Snouted Caiman (Caiman latirostris)Marcio AraujoNo ratings yet
- Activity 2 Biochemical Process Written Report - ArabDocument5 pagesActivity 2 Biochemical Process Written Report - Arabtrisha estrellaNo ratings yet
- Bahasa Inggris Iii: 4. Checking Vital SignDocument14 pagesBahasa Inggris Iii: 4. Checking Vital SignErlangga PratamaNo ratings yet
- Evidences of Evolution Activity SheetsDocument8 pagesEvidences of Evolution Activity SheetsVena Grace MendezNo ratings yet
- Pulmonary Ventilation ModuleDocument5 pagesPulmonary Ventilation ModulejunquelalaNo ratings yet
- Insect Ecology (Fourth Edition), 2016 - Timothy SchowalterDocument12 pagesInsect Ecology (Fourth Edition), 2016 - Timothy SchowalterJefferson SepúlvedaNo ratings yet
- Muscular System Chapter 10 SummaryDocument6 pagesMuscular System Chapter 10 SummaryKYLA LORENZ BALUMANo ratings yet
- Anatomy and Physiology MnemonicsDocument7 pagesAnatomy and Physiology MnemonicsLalajimNo ratings yet
- Biological OxidationDocument114 pagesBiological OxidationvanshikaNo ratings yet
- External Anatomy of Toad: Stephen C. Capilitan, MSTDocument24 pagesExternal Anatomy of Toad: Stephen C. Capilitan, MSTSofia RatuisteNo ratings yet
- Theory and Strategy in HistochemistryDocument581 pagesTheory and Strategy in HistochemistryNoa GoyanesNo ratings yet
- Lipids Chem MBBSDocument57 pagesLipids Chem MBBSUmar JibrinNo ratings yet
- Aerobic Respiration 1 QPDocument13 pagesAerobic Respiration 1 QPKHANSANo ratings yet
- The Meaning of My GemstonesDocument10 pagesThe Meaning of My GemstonesAngelBaby38No ratings yet
- Physiology RAASDocument3 pagesPhysiology RAASAulia Mahya FaradisaNo ratings yet
- EosinophiliaDocument8 pagesEosinophiliajohnNo ratings yet
- Nutritional Support in The Perioperative Period: 4 Fluid Balance and Metabolism in SurgeryDocument29 pagesNutritional Support in The Perioperative Period: 4 Fluid Balance and Metabolism in SurgeryFreddy Negron HuamaniNo ratings yet
- Herpetology: An Introductory Biology of Amphibians and ReptilesDocument4 pagesHerpetology: An Introductory Biology of Amphibians and ReptilesChristine BantilingNo ratings yet
- Platelet Rich Plasma, A Powerful Tool in DermatologyDocument10 pagesPlatelet Rich Plasma, A Powerful Tool in DermatologyJulian SilitongaNo ratings yet
- The Interaction Between Autophagy, Helicobacter Pylori, and Gut Microbiota in Gastric CarcinogenesisDocument20 pagesThe Interaction Between Autophagy, Helicobacter Pylori, and Gut Microbiota in Gastric CarcinogenesisWaldo IzyNo ratings yet
- Regulation of Uterine Function: A Biochemical Conundrum in The Regulation of Smooth Muscle RelaxationDocument9 pagesRegulation of Uterine Function: A Biochemical Conundrum in The Regulation of Smooth Muscle Relaxationtiga sekawan sekawanNo ratings yet
- NCP For Diabetic KetoacidosisDocument2 pagesNCP For Diabetic KetoacidosisLovely Cacapit100% (1)
- Animal Structure and FunctionDocument49 pagesAnimal Structure and FunctionJohn Steven AalaNo ratings yet
- Neurotransmitter Dalam Fisiologi Saraf Otonom: Iwan Dwi Cahyono, Himawan Sasongko, Aria Dian PrimatikaDocument14 pagesNeurotransmitter Dalam Fisiologi Saraf Otonom: Iwan Dwi Cahyono, Himawan Sasongko, Aria Dian PrimatikaardianNo ratings yet
- Thermoregulation: Presenter: Dr. Faisal Djamil Sheriff, H58/38054/2020 Moderator: Dr. MuriithiDocument56 pagesThermoregulation: Presenter: Dr. Faisal Djamil Sheriff, H58/38054/2020 Moderator: Dr. MuriithiHarshilPatelNo ratings yet
- Central Nervous System Tumors: GeneralDocument35 pagesCentral Nervous System Tumors: GeneralleartaNo ratings yet
- Understanding Insect Life CyclesDocument38 pagesUnderstanding Insect Life CyclesYeye bonnellNo ratings yet
- VETRO DESIGN SRL Medical CatalogDocument104 pagesVETRO DESIGN SRL Medical CatalogCristiDanNo ratings yet
- 8 Inch StickDocument29 pages8 Inch Stickworkbeast24No ratings yet
- Final OsmosisDocument2 pagesFinal Osmosisapi-523278684No ratings yet
- Study of Cancer: By: Parth Mishra XII-ScienceDocument30 pagesStudy of Cancer: By: Parth Mishra XII-ScienceParth Mishra100% (1)