You might also like
- Microbiome and Metabolome in Diagnosis, Therapy, and other Strategic ApplicationsFrom EverandMicrobiome and Metabolome in Diagnosis, Therapy, and other Strategic ApplicationsJoel FaintuchNo ratings yet
- The Bowman-Birk Inhibitor From Soybeans As An Anticarcinogenic AgentDocument7 pagesThe Bowman-Birk Inhibitor From Soybeans As An Anticarcinogenic AgentTalhaNo ratings yet
- Biotin and Other Interferences in Immunoassays: A Concise GuideFrom EverandBiotin and Other Interferences in Immunoassays: A Concise GuideNo ratings yet
- Medicines 05 00101Document14 pagesMedicines 05 00101Alice de MoraesNo ratings yet
- Pre 2Document6 pagesPre 2ruNo ratings yet
- Probiotik Mengurangi Resistensi AntibiotikDocument34 pagesProbiotik Mengurangi Resistensi AntibiotikviaNo ratings yet
- A Review of The Effects of Natural Compounds, Medical Plants, Mushrooms On Thegut Microbiota and Colitis and CancerDocument8 pagesA Review of The Effects of Natural Compounds, Medical Plants, Mushrooms On Thegut Microbiota and Colitis and CancerMartinaNo ratings yet
- Desordenes Gastrointestinales PDFDocument7 pagesDesordenes Gastrointestinales PDFMvz Mary Cielo Fernández FuentesNo ratings yet
- Bacteriocins and Their Food ApplicationsDocument19 pagesBacteriocins and Their Food Applicationsanon_206605756No ratings yet
- Diet Composition and Non-Antibiotic Feeding Programs in EuropeDocument14 pagesDiet Composition and Non-Antibiotic Feeding Programs in EuropeDalia MatjodNo ratings yet
- AAFP 2017 - Probiotics For Gastrointestinal Conditions - A Summary of EvidenceDocument10 pagesAAFP 2017 - Probiotics For Gastrointestinal Conditions - A Summary of EvidenceBigPharma HealtcareNo ratings yet
- Using Probioticos in Gastrointestinal DisordersDocument7 pagesUsing Probioticos in Gastrointestinal DisordersRonald CampoverdeNo ratings yet
- Comparative Evaluation of The Anti-Angiogenic Properties of Vitamin B Complex and Ascorbic Acid Using Duck Embryo ChorioallaDocument7 pagesComparative Evaluation of The Anti-Angiogenic Properties of Vitamin B Complex and Ascorbic Acid Using Duck Embryo ChorioallaIOER International Multidisciplinary Research Journal ( IIMRJ)No ratings yet
- Gut Microbiome and Breast Cancer MetastasisDocument4 pagesGut Microbiome and Breast Cancer Metastasiserdost450No ratings yet
- Nutrients 13 00526Document16 pagesNutrients 13 00526yabaNo ratings yet
- Advances in Cancer Biology - MetastasisDocument12 pagesAdvances in Cancer Biology - MetastasisMuhamad AliNo ratings yet
- Modulation of The Gastrointestinal Microbiome With Nondigestible Fermentable Carbohydrates To Improve Human HealthDocument35 pagesModulation of The Gastrointestinal Microbiome With Nondigestible Fermentable Carbohydrates To Improve Human HealthslothwonNo ratings yet
- Focus4 Jun 2020Document6 pagesFocus4 Jun 2020Ahmed HamdyNo ratings yet
- NutrientsDocument13 pagesNutrientsDyane VatriciaNo ratings yet
- Using Bifidobacterium and Propionibacterium Strains in Probiotic Consortia To Normalize The Gastrointestinal TractDocument17 pagesUsing Bifidobacterium and Propionibacterium Strains in Probiotic Consortia To Normalize The Gastrointestinal TractThiago MalpighiNo ratings yet
- Intestinal Colonisation, Microbiota and Future Probiotics?: Original ArticleDocument5 pagesIntestinal Colonisation, Microbiota and Future Probiotics?: Original ArticleSherif Abou BakrNo ratings yet
- Efficacy of Bacillus Probiotics in Prevention of Antibiotic-Associated Diarrhoea: A Randomized, Double-Blind, Placebo-Controlled Clinical TrialDocument6 pagesEfficacy of Bacillus Probiotics in Prevention of Antibiotic-Associated Diarrhoea: A Randomized, Double-Blind, Placebo-Controlled Clinical Trialdrfaruqui2551No ratings yet
- 1 s2.0 S1756464620303042 MainDocument12 pages1 s2.0 S1756464620303042 MainValentina IorgaNo ratings yet
- Clinical Uses of Probiotics: S R M - ADocument5 pagesClinical Uses of Probiotics: S R M - AWahyu RedfieldNo ratings yet
- Food Research InternationalDocument10 pagesFood Research InternationalSandraJSantosNo ratings yet
- Artigo Pics 4 PDFDocument9 pagesArtigo Pics 4 PDFGabriela Brito de Lima SilvaNo ratings yet
- Microbiome in Gastrointestinal Cancer 2023Document299 pagesMicrobiome in Gastrointestinal Cancer 2023xtz9bcqm7sNo ratings yet
- Chemopreventive Effect of Dietary Anthocyanins Against Gastrointestinal Cancers: A Review of Recent Advances and PerspectivesDocument2 pagesChemopreventive Effect of Dietary Anthocyanins Against Gastrointestinal Cancers: A Review of Recent Advances and PerspectivesNina Klairea PaduaNo ratings yet
- Xie PDocument19 pagesXie PSreeja CherukuruNo ratings yet
- Journal of The Saudi Society of Agricultural SciencesDocument8 pagesJournal of The Saudi Society of Agricultural SciencesNABILA JUNISKY SUSETYAWATINo ratings yet
- BiavatiB Probiotics and PrebioticsDocument75 pagesBiavatiB Probiotics and PrebioticsRashmin ShahNo ratings yet
- Probiotics Article 2019 PDFDocument10 pagesProbiotics Article 2019 PDFLuiis LimaNo ratings yet
- Engineering Probiotics For Therapeutic ApplicationsDocument9 pagesEngineering Probiotics For Therapeutic ApplicationsElinaNo ratings yet
- Probiotics - Potential Pharmaceutical Applications (2002) PDFDocument9 pagesProbiotics - Potential Pharmaceutical Applications (2002) PDFmilu1312No ratings yet
- The Benefits of Gene TherapyDocument3 pagesThe Benefits of Gene TherapyLevi AckermanNo ratings yet
- Probiotics Nutritional Therapeutic Tool 2329 8901 1000194Document8 pagesProbiotics Nutritional Therapeutic Tool 2329 8901 1000194Javier MartinNo ratings yet
- ByoticsDocument15 pagesByoticsEvelina VladNo ratings yet
- Cumin Prevents 17-Estradiol-Associated Breast Cancer in ACI Rats - Ijms-22-06194Document17 pagesCumin Prevents 17-Estradiol-Associated Breast Cancer in ACI Rats - Ijms-22-06194Teresa BeckNo ratings yet
- Colorektal CancerDocument24 pagesColorektal CancerAgus PraktyasaNo ratings yet
- Probiotics - Future LandscapesDocument16 pagesProbiotics - Future LandscapesJavier MartinNo ratings yet
- Microbiome and CancerDocument63 pagesMicrobiome and CancerK C Goutham ReddyNo ratings yet
- Cancers 11 00038Document26 pagesCancers 11 00038JK NUTRISIERVICENo ratings yet
- The Role of Vitamins in CancerDocument17 pagesThe Role of Vitamins in CancerNadir KoçakNo ratings yet
- MainDocument17 pagesMainMohammed KhalidNo ratings yet
- Articulo ProbioticosDocument13 pagesArticulo ProbioticososcarandresNo ratings yet
- Benjamin Bartsch, Chee Kin Then, Elinor Harriss, Christiana Kartsonaki, Anne E. KiltieDocument9 pagesBenjamin Bartsch, Chee Kin Then, Elinor Harriss, Christiana Kartsonaki, Anne E. KiltieChrisYapNo ratings yet
- Nihms 833860Document20 pagesNihms 833860yabaNo ratings yet
- Example 7Document5 pagesExample 7Tutor BlairNo ratings yet
- Gut Microbiota For Health Summit Report 2016Document4 pagesGut Microbiota For Health Summit Report 2016GMFHNo ratings yet
- BiosimilarsDocument13 pagesBiosimilarsAhmed Abd El Aziz100% (1)
- Bomba A Et Al 2002 Probiotics and PUFAsDocument5 pagesBomba A Et Al 2002 Probiotics and PUFAsSandip PatilNo ratings yet
- Probiotics and Health: A Review of The Evidence: E. WeichselbaumDocument35 pagesProbiotics and Health: A Review of The Evidence: E. WeichselbaumJavierjongNo ratings yet
- Nihms 1028791Document31 pagesNihms 1028791yabaNo ratings yet
- 1 s2.0 S0022316622004588 MainDocument11 pages1 s2.0 S0022316622004588 MainNejc KovačNo ratings yet
- Fungal Beta-Glucans As Adjuvants For Treating Cancer Patients - Revisão Sistemática de ECRDocument10 pagesFungal Beta-Glucans As Adjuvants For Treating Cancer Patients - Revisão Sistemática de ECRMaria Vitória Cota de AbreuNo ratings yet
- Pro y PrebioticosDocument21 pagesPro y PrebioticosIvan CLNo ratings yet
- Probiotics - Friendly Bacteria With A Host of Benefits: HistoryDocument4 pagesProbiotics - Friendly Bacteria With A Host of Benefits: Historynikunj_patel_81No ratings yet
- A Vegan Diet Alters The Human Colonic Faecal MicrobiotaDocument8 pagesA Vegan Diet Alters The Human Colonic Faecal MicrobiotaWilver Edson Otalora AnturianoNo ratings yet
- Recent Development of Probiotic Bifidobacteria For Treating Human DiseasesDocument17 pagesRecent Development of Probiotic Bifidobacteria For Treating Human DiseasesLinh HoangNo ratings yet
- Antitumor Effect of PyridoxineDocument4 pagesAntitumor Effect of PyridoxineNarcis IftimescuNo ratings yet
- Manila Standard Today - July 21, 2012 IssueDocument16 pagesManila Standard Today - July 21, 2012 IssueManila Standard TodayNo ratings yet
- 2.6. Curved Mirrors Calculations & Applications LessonDocument32 pages2.6. Curved Mirrors Calculations & Applications Lesson6h4gxcf7jbNo ratings yet
- (Routledge Research in Intellectual Property) Olasupo Ayodeji Owoeye - Intellectual Property and Access To Medicines in Africa - A Regional Framework For Access-Routledge (2019)Document291 pages(Routledge Research in Intellectual Property) Olasupo Ayodeji Owoeye - Intellectual Property and Access To Medicines in Africa - A Regional Framework For Access-Routledge (2019)Rajnia RodriguesNo ratings yet
- Urinary System and Disorders IntroductionDocument23 pagesUrinary System and Disorders IntroductionHananya ManroeNo ratings yet
- Annotated BibliographyDocument3 pagesAnnotated Bibliographyapi-272471717No ratings yet
- The Blood of DraculaDocument20 pagesThe Blood of DraculaJackSiedschlagNo ratings yet
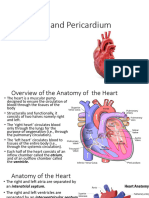
- The Heart and PericardiumDocument40 pagesThe Heart and PericardiumMartha Orendu Oche AttahNo ratings yet
- Lifespan: Why We Age - and Why We Don't Have To - DR David A. SinclairDocument5 pagesLifespan: Why We Age - and Why We Don't Have To - DR David A. Sinclairtidasesu0% (1)
- Clinical Study: Overjet and Overbite Influence On Cyclic Masticatory Movements: A CT StudyDocument7 pagesClinical Study: Overjet and Overbite Influence On Cyclic Masticatory Movements: A CT Studyarya091193No ratings yet
- 09 Marijuana Facts For TeensDocument24 pages09 Marijuana Facts For Teensapi-309082881No ratings yet
- Cases Journal: Acquired Nonobstructive Urinary Bladder Diverticulum: A CaseDocument3 pagesCases Journal: Acquired Nonobstructive Urinary Bladder Diverticulum: A CaseIrma Suriani DarwisNo ratings yet
- Chapter 4 Factors Affecting Performance: Training Purpose OnlyDocument19 pagesChapter 4 Factors Affecting Performance: Training Purpose OnlySajjad ShamimNo ratings yet
- Neurology II 6.04 Traumatic Brain Injury Dr. TanDocument10 pagesNeurology II 6.04 Traumatic Brain Injury Dr. TanAbi IgsNo ratings yet
- OSHA 3162 - Screening and SurveillanceDocument40 pagesOSHA 3162 - Screening and SurveillanceWahed Mn ElnasNo ratings yet
- Headache and Joint PainDocument13 pagesHeadache and Joint PainJosiah Noella BrizNo ratings yet
- Let's Talk Game CardsDocument4 pagesLet's Talk Game CardssycagurlNo ratings yet
- Journal Neuropediatri PDFDocument8 pagesJournal Neuropediatri PDFHalimah PramudiyantiNo ratings yet
- Unit 7 K10 Global SuccessDocument5 pagesUnit 7 K10 Global SuccessLý Nguyễn Hạnh NhungNo ratings yet
- NY B19 Misc FDR - WTC Disaster Response - NYCDOH - 9-12-01Document87 pagesNY B19 Misc FDR - WTC Disaster Response - NYCDOH - 9-12-019/11 Document Archive100% (2)
- Artificial IntelligenceDocument27 pagesArtificial IntelligenceMd HassanNo ratings yet
- Laboratory Procedure Protocols For Parasitological Sample Collection ProcessingDocument36 pagesLaboratory Procedure Protocols For Parasitological Sample Collection ProcessingDr.Kedar Karki ,M.V.Sc.Preventive Vet.Medicine CLSU Philippines100% (4)
- Toxoplasma GondiiDocument7 pagesToxoplasma GondiiDedy SavradinataNo ratings yet
- Language and The BrainDocument8 pagesLanguage and The BrainAntonio GoqueNo ratings yet
- Natiomal Plan of ActionDocument36 pagesNatiomal Plan of Actiontendai precious nyamadzaoNo ratings yet
- Npte Exam TablesDocument586 pagesNpte Exam TablesarbazNo ratings yet
- Tapping Into Body Joy PDFDocument18 pagesTapping Into Body Joy PDFKaren McLaughlin100% (13)
- Nursing Skills (Vital Signs) PDFDocument48 pagesNursing Skills (Vital Signs) PDFkhizer hayatNo ratings yet
- Types, Characteristics, and Identification of Learners With Difficulty Seeing - 0Document13 pagesTypes, Characteristics, and Identification of Learners With Difficulty Seeing - 0Laleth Mendoza OjalesNo ratings yet
- SSOPDocument133 pagesSSOPYanuar Rufianti Wongi100% (1)
- Love Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)From EverandLove Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)Rating: 3 out of 5 stars3/5 (1)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 4.5 out of 5 stars4.5/5 (82)
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (32)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (404)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDFrom EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDRating: 5 out of 5 stars5/5 (3)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 4 out of 5 stars4/5 (4)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeRating: 2 out of 5 stars2/5 (1)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (42)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeFrom EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeRating: 4.5 out of 5 stars4.5/5 (254)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (170)
- Manipulation: The Ultimate Guide To Influence People with Persuasion, Mind Control and NLP With Highly Effective Manipulation TechniquesFrom EverandManipulation: The Ultimate Guide To Influence People with Persuasion, Mind Control and NLP With Highly Effective Manipulation TechniquesRating: 4.5 out of 5 stars4.5/5 (1412)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaFrom EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaRating: 4.5 out of 5 stars4.5/5 (266)
- To Explain the World: The Discovery of Modern ScienceFrom EverandTo Explain the World: The Discovery of Modern ScienceRating: 3.5 out of 5 stars3.5/5 (51)
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 4 out of 5 stars4/5 (6)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryFrom EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryRating: 4 out of 5 stars4/5 (46)
- The Marshmallow Test: Mastering Self-ControlFrom EverandThe Marshmallow Test: Mastering Self-ControlRating: 4.5 out of 5 stars4.5/5 (60)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (328)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4.5 out of 5 stars4.5/5 (6)
- How to ADHD: The Ultimate Guide and Strategies for Productivity and Well-BeingFrom EverandHow to ADHD: The Ultimate Guide and Strategies for Productivity and Well-BeingRating: 1 out of 5 stars1/5 (1)