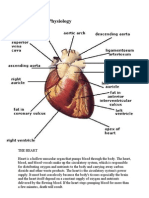
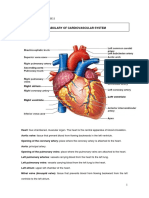
You might also like
- Heart: StructureDocument11 pagesHeart: StructureKert trocioNo ratings yet
- Circulatory System: A Tutorial Study GuideFrom EverandCirculatory System: A Tutorial Study GuideRating: 5 out of 5 stars5/5 (3)
- Cardiovascular Physiology - Dra. ValerioDocument16 pagesCardiovascular Physiology - Dra. ValerioAlexandra Duque-David100% (2)
- Coronary Heart Disease: A Guide to Diagnosis and TreatmentFrom EverandCoronary Heart Disease: A Guide to Diagnosis and TreatmentRating: 3.5 out of 5 stars3.5/5 (3)
- Cardiovascular SystemDocument32 pagesCardiovascular SystemAugustus Alejandro ZenitNo ratings yet
- Anatomy Physiology HeartDocument9 pagesAnatomy Physiology HeartAlvheen JoaquinNo ratings yet
- Human Body Book | Introduction to the Vascular System | Children's Anatomy & Physiology EditionFrom EverandHuman Body Book | Introduction to the Vascular System | Children's Anatomy & Physiology EditionNo ratings yet
- Anatomy Assessment 12 (AutoRecovered)Document7 pagesAnatomy Assessment 12 (AutoRecovered)Jellisa Fuentes AbrahamNo ratings yet
- Anatomy Assessment 12 (AutoRecovered)Document7 pagesAnatomy Assessment 12 (AutoRecovered)Jellisa Fuentes AbrahamNo ratings yet
- CNCQ Basic Reading Chapter 7Document10 pagesCNCQ Basic Reading Chapter 7Thiện NguyễnNo ratings yet
- Anatomy and Physiology of The HeartDocument3 pagesAnatomy and Physiology of The HeartjeffcaselNo ratings yet
- Cardio Vascular SystemDocument8 pagesCardio Vascular SystemguptaasitNo ratings yet
- Vocabulary of Cardiovascular System: Ingles Medico Ii - Clase 2Document7 pagesVocabulary of Cardiovascular System: Ingles Medico Ii - Clase 2Agustín Bravo Arreyes100% (2)
- CHAPTER 3 GcsDocument14 pagesCHAPTER 3 GcsNurse NotesNo ratings yet
- Anatomy and Physiology of the Heart ExplainedDocument6 pagesAnatomy and Physiology of the Heart ExplainedChamCham AquinoNo ratings yet
- Project BioDocument18 pagesProject Bioyanshu falduNo ratings yet
- HUMAN ANATOMY AND PHYSIOLOGY LABORATORY Cardiovascular SystemDocument12 pagesHUMAN ANATOMY AND PHYSIOLOGY LABORATORY Cardiovascular SystemZaira MangalimanNo ratings yet
- Coronary Artery Disease - Case StudyDocument19 pagesCoronary Artery Disease - Case StudyJulieteeySarania100% (3)
- The HeartDocument11 pagesThe HeartElisa TapalesNo ratings yet
- Chapter 13 - HeartDocument5 pagesChapter 13 - Heartbussniesm05No ratings yet
- Heart and BrainDocument29 pagesHeart and BrainDIPANJAN MAJUMDAR100% (6)
- Heart Anatomy ExplainedDocument15 pagesHeart Anatomy ExplainedAritra SahaNo ratings yet
- Heart Structure, Function and Arrhythmias: Elizabeth M. Cherry and Flavio H. FentonDocument15 pagesHeart Structure, Function and Arrhythmias: Elizabeth M. Cherry and Flavio H. Fentonmaurice bertrandNo ratings yet
- Our Heart BeatsDocument7 pagesOur Heart BeatsferyNo ratings yet
- Anatomy & PhysiologyDocument57 pagesAnatomy & PhysiologyBrittanie MouzonNo ratings yet
- Cardiovascular System: Halabja Technical College of Applied SciencesDocument6 pagesCardiovascular System: Halabja Technical College of Applied SciencesELYANNo ratings yet
- Cardiovascular SystemDocument9 pagesCardiovascular SystemWild RiftNo ratings yet
- Chapter VIII CardiovascularDocument7 pagesChapter VIII CardiovascularTitoMacoyTVNo ratings yet
- Anatomy and Physiology Lecture NotesDocument6 pagesAnatomy and Physiology Lecture NotesBlackcat KememeyNo ratings yet
- Anatomy and Physiology of The HeartDocument8 pagesAnatomy and Physiology of The HeartJoan Devera100% (1)
- Circulatory SystemDocument111 pagesCirculatory SystemYounas Bhatti100% (1)
- The Cardiovascular System The HeartDocument9 pagesThe Cardiovascular System The HeartMarjorie AbelgasNo ratings yet
- The HeartDocument24 pagesThe HeartAnubhav MishraNo ratings yet
- The Circulatory System: Transport for CellsDocument4 pagesThe Circulatory System: Transport for CellsPuguh HadiNo ratings yet
- The Circulatory System: How Blood Flows Through the Heart & BodyDocument3 pagesThe Circulatory System: How Blood Flows Through the Heart & BodyNinaNo ratings yet
- A Case Study About Cardiovascular DiseaseDocument10 pagesA Case Study About Cardiovascular DiseaseBarben Balaquit100% (1)
- Cardiac Control CentreDocument8 pagesCardiac Control CentregeorgeloxdaleNo ratings yet
- Anatomy and Physiology of the HeartDocument4 pagesAnatomy and Physiology of the HeartnoviantyNo ratings yet
- Anatomy Lecture Notes Unit 7 Circulatory System - The HeartDocument7 pagesAnatomy Lecture Notes Unit 7 Circulatory System - The Heartrapunzel777No ratings yet
- The Circulatory System: Agriscience 332 Animal Science #8646-A TEKS: (C) (2) (A) and (C) (2) (B)Document111 pagesThe Circulatory System: Agriscience 332 Animal Science #8646-A TEKS: (C) (2) (A) and (C) (2) (B)brdaisyNo ratings yet
- Oral RevDocument6 pagesOral RevjjolaguerNo ratings yet
- CardiovascularDocument30 pagesCardiovascularBeverly LavillaNo ratings yet
- Anatomy of The HeartDocument5 pagesAnatomy of The HeartxXjhmXxNo ratings yet
- CardiovascularDocument30 pagesCardiovascularKristine Joy Librando EscalanteNo ratings yet
- Anatomy of Heart - RyanDocument5 pagesAnatomy of Heart - Ryandrowned_clarkeNo ratings yet
- Structure and Function of the HeartDocument5 pagesStructure and Function of the Heartdrowned_clarkeNo ratings yet
- Anatomy and Physiology of the HeartDocument8 pagesAnatomy and Physiology of the HeartAbigail BrillantesNo ratings yet
- Management of Clients With Problems of The Cardiovascular SystemDocument7 pagesManagement of Clients With Problems of The Cardiovascular Systemjerry smatt dumtoNo ratings yet
- Cardiovascular SystemDocument29 pagesCardiovascular Systemethel roseNo ratings yet
- Assignment On Anatomy and Physiology of Heart: Submitted By: Submitted To: Jayita Gayen Madam S. PoddarDocument10 pagesAssignment On Anatomy and Physiology of Heart: Submitted By: Submitted To: Jayita Gayen Madam S. PoddarJayita Gayen DuttaNo ratings yet
- Structure and Function of the HeartDocument5 pagesStructure and Function of the Heartdrowned_clarkeNo ratings yet
- Function Human HeartDocument13 pagesFunction Human Heartshyam_inkNo ratings yet
- Chapter I - Cardiovascular System: HeartDocument2 pagesChapter I - Cardiovascular System: HeartIndranil SinhaNo ratings yet
- MS Wrep IIDocument51 pagesMS Wrep IIiana-almocera-6970No ratings yet
- W.G. Igcse Transport in HumansDocument20 pagesW.G. Igcse Transport in Humansbloodshooter1996No ratings yet
- The Cardiovascular System: Arteries VeinsDocument27 pagesThe Cardiovascular System: Arteries VeinsMark Anthony DiegoNo ratings yet
- Anatomy of Cardiovascular SystemDocument23 pagesAnatomy of Cardiovascular SystemElvira Wisakhadewi100% (1)
- Cardiovascular SystemDocument7 pagesCardiovascular SystemAbdallah AlasalNo ratings yet
- French Number System. LapuzDocument2 pagesFrench Number System. LapuzLiam Jacque LapuzNo ratings yet
- S-BIOL121 BSY32 1st Sem (2022-2023) - Https - Dlsud - Edu20.org - Student - Quiz - Assignment - Submissions - 36301681Document3 pagesS-BIOL121 BSY32 1st Sem (2022-2023) - Https - Dlsud - Edu20.org - Student - Quiz - Assignment - Submissions - 36301681Liam Jacque LapuzNo ratings yet
- Frog respiratory and urogenital systems worksheetDocument8 pagesFrog respiratory and urogenital systems worksheetLiam Jacque LapuzNo ratings yet
- Auditing Theory - WILEYDocument21 pagesAuditing Theory - WILEYCyd Chary Limbaga BiadnesNo ratings yet
- S-BIOL121 BSY32 1st Sem (2022-2023) - Https - Dlsud - Edu20.org - Student - Take - Quiz - Assignment - Start - 36301681Document4 pagesS-BIOL121 BSY32 1st Sem (2022-2023) - Https - Dlsud - Edu20.org - Student - Take - Quiz - Assignment - Start - 36301681Liam Jacque LapuzNo ratings yet
- MusclesssDocument4 pagesMusclesssLiam Jacque LapuzNo ratings yet
- The Effect of Font Weight On Reading Speed and Working Memory Recall Grp. 4 BSY32Document51 pagesThe Effect of Font Weight On Reading Speed and Working Memory Recall Grp. 4 BSY32Liam Jacque LapuzNo ratings yet
- Cardiovascular System Lesson on Blood Vessels, Blood PressureDocument5 pagesCardiovascular System Lesson on Blood Vessels, Blood PressureLiam Jacque LapuzNo ratings yet
- Lesson 8. Holistic Dynamic Theory by MaslowDocument8 pagesLesson 8. Holistic Dynamic Theory by MaslowLiam Jacque LapuzNo ratings yet
- Skinner's Behavioral Analysis TheoryDocument7 pagesSkinner's Behavioral Analysis TheoryLiam Jacque LapuzNo ratings yet
- Amino Acid Titration. Sci PaperDocument5 pagesAmino Acid Titration. Sci PaperLiam Jacque LapuzNo ratings yet
- Lesson 6 Humanistic Psychoanalysis: C-Psym213 Theories of Personality Module 5: Humanistic Psychoanalysis by FrommDocument4 pagesLesson 6 Humanistic Psychoanalysis: C-Psym213 Theories of Personality Module 5: Humanistic Psychoanalysis by FrommLiam Jacque LapuzNo ratings yet
- FGD in Cebu: HIV, Gender-based Violence, and DiscriminationDocument2 pagesFGD in Cebu: HIV, Gender-based Violence, and DiscriminationLiam Jacque LapuzNo ratings yet
- Bone AnatomyDocument11 pagesBone AnatomyLiam Jacque LapuzNo ratings yet
- The General History of Virginia (1607-1614)Document10 pagesThe General History of Virginia (1607-1614)Liam Jacque LapuzNo ratings yet
- SDG 3 Final Output FinalsDocument3 pagesSDG 3 Final Output FinalsLiam Jacque LapuzNo ratings yet
- The General History of VirginiaDocument17 pagesThe General History of VirginiaLiam Jacque LapuzNo ratings yet
- General History of Virginia TextDocument5 pagesGeneral History of Virginia TextPamela BanayatNo ratings yet
- Colonialism: Colonialism Is A Practice of Domination, Which Involves The Subjugation of One People To Another"Document2 pagesColonialism: Colonialism Is A Practice of Domination, Which Involves The Subjugation of One People To Another"Liam Jacque LapuzNo ratings yet
- Characters Search Author: Six in of AnDocument8 pagesCharacters Search Author: Six in of AnLiam Jacque LapuzNo ratings yet
- PoetryDocument2 pagesPoetryLiam Jacque LapuzNo ratings yet
- Psychology Doctoral Program Experiences and Student Well-Being, Mental Health, and Optimism During The COVID-19 PandemicDocument11 pagesPsychology Doctoral Program Experiences and Student Well-Being, Mental Health, and Optimism During The COVID-19 PandemicLiam Jacque LapuzNo ratings yet
- Psychology Doctoral Program Experiences and Student Well-Being, Mental Health, and Optimism During The COVID-19 PandemicDocument11 pagesPsychology Doctoral Program Experiences and Student Well-Being, Mental Health, and Optimism During The COVID-19 PandemicLiam Jacque LapuzNo ratings yet
- Assessing Filipino Students' Readiness for E-LearningDocument12 pagesAssessing Filipino Students' Readiness for E-LearningLiam Jacque LapuzNo ratings yet
- Optimism and Well-Being in Times of COVID-19: Master ThesisDocument49 pagesOptimism and Well-Being in Times of COVID-19: Master ThesisLiam Jacque LapuzNo ratings yet
- W 27392Document26 pagesW 27392Red StohlNo ratings yet
- Efficacy, Hope, Optimism and Resilience at Workplace - Positive Organizational BehaviorDocument4 pagesEfficacy, Hope, Optimism and Resilience at Workplace - Positive Organizational BehaviorLiam Jacque LapuzNo ratings yet
- Assessing Filipino Students' Readiness for E-LearningDocument12 pagesAssessing Filipino Students' Readiness for E-LearningLiam Jacque LapuzNo ratings yet
- Fostering Networking Behavior Career PlaDocument11 pagesFostering Networking Behavior Career PlaLiam Jacque LapuzNo ratings yet
- MCQ Rognidan 170820Document58 pagesMCQ Rognidan 170820Setu UpadhyayNo ratings yet
- Dopamine Drug StudyDocument3 pagesDopamine Drug StudyNicole Soo78% (9)
- Chapter 4 (Cardiovascular Measurement) PDFDocument64 pagesChapter 4 (Cardiovascular Measurement) PDFFariza ZahariNo ratings yet
- Interpretasi EKGDocument81 pagesInterpretasi EKGGyna MarsianaNo ratings yet
- Overview of Cardiovascular System: An Introduction To Chapters 9 - 24 and Chapter 36 Guyton and Hall, 12 EditionDocument10 pagesOverview of Cardiovascular System: An Introduction To Chapters 9 - 24 and Chapter 36 Guyton and Hall, 12 EditionbahahahahNo ratings yet
- Multi-parameter Portable Patient Monitor DS 3000 Features and SpecsDocument2 pagesMulti-parameter Portable Patient Monitor DS 3000 Features and SpecsFrancisco TorresNo ratings yet
- Basic EKG Review: Identify Rhythms & InterventionsDocument31 pagesBasic EKG Review: Identify Rhythms & Interventionsmeb100% (1)
- January 2018 (IAL) QP - Unit 1 Edexcel Biology A-LevelDocument24 pagesJanuary 2018 (IAL) QP - Unit 1 Edexcel Biology A-LevelGhaziaNo ratings yet
- Heart BlockDocument20 pagesHeart BlockSumathi GopinathNo ratings yet
- Cardiology Guide: Key Concepts, Formulas and Treatment StrategiesDocument5 pagesCardiology Guide: Key Concepts, Formulas and Treatment StrategiesRodrigo FonsecaNo ratings yet
- Concise CardiologyDocument354 pagesConcise CardiologyAtish_Mathur_9874No ratings yet
- Dr. Tunggul - HHS2020Document27 pagesDr. Tunggul - HHS2020Riris SihotangNo ratings yet
- 1 Cardiovascular SystemDocument51 pages1 Cardiovascular SystemChryzl JoyNo ratings yet
- ANAT Unit 3 Cardiac Conduction System NotesDocument15 pagesANAT Unit 3 Cardiac Conduction System NotesSilvia-Mihaela TatomirNo ratings yet
- ACLS Cheat SheetDocument8 pagesACLS Cheat SheetLenTheRN85% (27)
- Hendra - EKGDocument34 pagesHendra - EKGRINANo ratings yet
- Acute & Chronic Heart Failure: Dr. Nurkhalis, SPJP, FihaDocument98 pagesAcute & Chronic Heart Failure: Dr. Nurkhalis, SPJP, FihablackcatNo ratings yet
- Cardiovascular Physiology: Cardiac Output, Blood Flow ControlsDocument33 pagesCardiovascular Physiology: Cardiac Output, Blood Flow ControlsLuiz Jorge MendonçaNo ratings yet
- The Circulatory SystemDocument8 pagesThe Circulatory SystemAymen MouradNo ratings yet
- Ecg Cheat Sheet 35Document2 pagesEcg Cheat Sheet 35jessjaylee80% (5)
- Acls MegacodeDocument8 pagesAcls MegacodeRachel PerandoNo ratings yet
- Chapter 15 - Vascular Distensibility and Function of Blood Vessels-Dr Moni Nader-13012019Document30 pagesChapter 15 - Vascular Distensibility and Function of Blood Vessels-Dr Moni Nader-13012019muna sNo ratings yet
- Ecg InterpretationDocument3 pagesEcg Interpretationman0billi0% (1)
- CardioDocument48 pagesCardiomalisalukmanNo ratings yet
- Artikel Gerontik CempakaDocument16 pagesArtikel Gerontik CempakaLatifah Nur JannahNo ratings yet
- January 2022 (IAL) QPDocument28 pagesJanuary 2022 (IAL) QPMaria Shabbir PanjiwalaNo ratings yet
- Circulatory System LabDocument3 pagesCirculatory System LabMatt Joseph CabantingNo ratings yet
- Cardiac EmergenciesDocument29 pagesCardiac EmergenciesAhmad MohamadNo ratings yet
- Dysrhythmias Nursing InterventionsDocument7 pagesDysrhythmias Nursing InterventionsGail Lian Santos100% (1)
- ACCSAP 10 Qs & As ReviewDocument470 pagesACCSAP 10 Qs & As ReviewAdeel Lakhiar100% (9)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (402)
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (14)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (78)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (169)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeNo ratings yet
- Techniques Exercises And Tricks For Memory ImprovementFrom EverandTechniques Exercises And Tricks For Memory ImprovementRating: 4.5 out of 5 stars4.5/5 (40)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisFrom EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (1)
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 5 out of 5 stars5/5 (4)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (42)
- The Ultimate Guide To Memory Improvement TechniquesFrom EverandThe Ultimate Guide To Memory Improvement TechniquesRating: 5 out of 5 stars5/5 (34)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingFrom EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingRating: 5 out of 5 stars5/5 (5)
- The Happiness Trap: How to Stop Struggling and Start LivingFrom EverandThe Happiness Trap: How to Stop Struggling and Start LivingRating: 4 out of 5 stars4/5 (1)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- The Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsFrom EverandThe Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsNo ratings yet
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (327)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeFrom EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeRating: 4.5 out of 5 stars4.5/5 (253)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingFrom EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingRating: 3.5 out of 5 stars3.5/5 (33)
- Daniel Kahneman's "Thinking Fast and Slow": A Macat AnalysisFrom EverandDaniel Kahneman's "Thinking Fast and Slow": A Macat AnalysisRating: 3.5 out of 5 stars3.5/5 (130)
- Summary: It Didn't Start with You: How Inherited Family Trauma Shapes Who We Are and How to End the Cycle By Mark Wolynn: Key Takeaways, Summary & AnalysisFrom EverandSummary: It Didn't Start with You: How Inherited Family Trauma Shapes Who We Are and How to End the Cycle By Mark Wolynn: Key Takeaways, Summary & AnalysisRating: 5 out of 5 stars5/5 (3)