You might also like
- Ecological Understanding: The Nature of Theory and the Theory of NatureFrom EverandEcological Understanding: The Nature of Theory and the Theory of NatureNo ratings yet
- Evolution and the Machinery of Chance: Philosophy, Probability, and Scientific Practice in BiologyFrom EverandEvolution and the Machinery of Chance: Philosophy, Probability, and Scientific Practice in BiologyNo ratings yet
- Making Sense of Evolution: The Conceptual Foundations of Evolutionary BiologyFrom EverandMaking Sense of Evolution: The Conceptual Foundations of Evolutionary BiologyRating: 4 out of 5 stars4/5 (3)
- The Third Lens: Metaphor and the Creation of Modern Cell BiologyFrom EverandThe Third Lens: Metaphor and the Creation of Modern Cell BiologyNo ratings yet
- The Evolution Controversy: A Survey of Competing TheoriesFrom EverandThe Evolution Controversy: A Survey of Competing TheoriesRating: 4.5 out of 5 stars4.5/5 (3)
- What Can History Tell Us About Founding Ethics On Biology?: Book ReviewDocument14 pagesWhat Can History Tell Us About Founding Ethics On Biology?: Book Reviewflora mauricioNo ratings yet
- Comments on “A Bio-Cultural-Historical Approach to the Study of Development (2016)”From EverandComments on “A Bio-Cultural-Historical Approach to the Study of Development (2016)”No ratings yet
- The New Biology: A Battle between Mechanism and OrganicismFrom EverandThe New Biology: A Battle between Mechanism and OrganicismNo ratings yet
- Fact and Method: Explanation, Confirmation and Reality in the Natural and the Social SciencesFrom EverandFact and Method: Explanation, Confirmation and Reality in the Natural and the Social SciencesRating: 3 out of 5 stars3/5 (2)
- Articulating the World: Conceptual Understanding and the Scientific ImageFrom EverandArticulating the World: Conceptual Understanding and the Scientific ImageRating: 5 out of 5 stars5/5 (1)
- Darwinian Reductionism: Or, How to Stop Worrying and Love Molecular BiologyFrom EverandDarwinian Reductionism: Or, How to Stop Worrying and Love Molecular BiologyRating: 4 out of 5 stars4/5 (4)
- Biological Individuality: Integrating Scientific, Philosophical, and Historical PerspectivesFrom EverandBiological Individuality: Integrating Scientific, Philosophical, and Historical PerspectivesScott LidgardRating: 2 out of 5 stars2/5 (1)
- The Welcomed Tension Between Philosophy and ScienceDocument12 pagesThe Welcomed Tension Between Philosophy and ScienceDelfin CalderolliNo ratings yet
- The Theory of Evolution: Principles, Concepts, and AssumptionsFrom EverandThe Theory of Evolution: Principles, Concepts, and AssumptionsNo ratings yet
- Populations in a Seasonal Environment. (MPB-5)From EverandPopulations in a Seasonal Environment. (MPB-5)Rating: 5 out of 5 stars5/5 (1)
- A Hierarchical Concept of Ecosystems. (MPB-23), Volume 23From EverandA Hierarchical Concept of Ecosystems. (MPB-23), Volume 23No ratings yet
- Social Practices as Biological Niche ConstructionFrom EverandSocial Practices as Biological Niche ConstructionRating: 5 out of 5 stars5/5 (1)
- Keller, E. F. - A Clash of Two Cultures (2007)Document1 pageKeller, E. F. - A Clash of Two Cultures (2007)ronaldbeatNo ratings yet
- Articulo 5 PDFDocument11 pagesArticulo 5 PDFangiw fuentesNo ratings yet
- Scientific Argumentation in Biology: 30 Classroom ActivitiesFrom EverandScientific Argumentation in Biology: 30 Classroom ActivitiesNo ratings yet
- Groupthink in Science: Greed, Pathological Altruism, Ideology, Competition, and CultureFrom EverandGroupthink in Science: Greed, Pathological Altruism, Ideology, Competition, and CultureNo ratings yet
- Systems Theory and Theology: The Living Interplay between Science and ReligionFrom EverandSystems Theory and Theology: The Living Interplay between Science and ReligionNo ratings yet
- Philosophy of Science An Analysis of QuaDocument10 pagesPhilosophy of Science An Analysis of Quayasmim paixãoNo ratings yet
- Dreamers, Visionaries, and Revolutionaries in the Life SciencesFrom EverandDreamers, Visionaries, and Revolutionaries in the Life SciencesNo ratings yet
- Sex and Death: An Introduction to Philosophy of BiologyFrom EverandSex and Death: An Introduction to Philosophy of BiologyRating: 3.5 out of 5 stars3.5/5 (10)
- Science's Blind Spot: The Unseen Religion of Scientific NaturalismFrom EverandScience's Blind Spot: The Unseen Religion of Scientific NaturalismRating: 4.5 out of 5 stars4.5/5 (3)
- Caplan TestabilityDisreputabilityStructure 1978Document19 pagesCaplan TestabilityDisreputabilityStructure 1978marvasNo ratings yet
- The Whole Truth: A Cosmologist’s Reflections on the Search for Objective RealityFrom EverandThe Whole Truth: A Cosmologist’s Reflections on the Search for Objective RealityNo ratings yet
- Sayonara Theory of Evolution, Hello Biological InterrelationFrom EverandSayonara Theory of Evolution, Hello Biological InterrelationNo ratings yet
- The Common Extremalities in Biology and Physics: Maximum Energy Dissipation Principle in Chemistry, Biology, Physics and EvolutionFrom EverandThe Common Extremalities in Biology and Physics: Maximum Energy Dissipation Principle in Chemistry, Biology, Physics and EvolutionNo ratings yet
- One World: The Interaction of Science and TheologyFrom EverandOne World: The Interaction of Science and TheologyRating: 3.5 out of 5 stars3.5/5 (5)
- Donoghue, 1985. A Critique of The Biological Species Concept and Recommendations For A Phylogenetic AlternativeDocument11 pagesDonoghue, 1985. A Critique of The Biological Species Concept and Recommendations For A Phylogenetic AlternativeRaissa SiqueiraNo ratings yet
- (Cambridge Introductions To Philosophy) Kent W. Staley - An Introduction To The Philosophy of Science-Cambridge University Press (2014) PDFDocument212 pages(Cambridge Introductions To Philosophy) Kent W. Staley - An Introduction To The Philosophy of Science-Cambridge University Press (2014) PDFRianda Marta Derici100% (5)
- Do We Need A Scientific Revolution?: WWW - Nick-Maxwell - Demon.co - UkDocument16 pagesDo We Need A Scientific Revolution?: WWW - Nick-Maxwell - Demon.co - UkAngel Tabuena IINo ratings yet
- Assignment 2: Module Code: HN5001 Module Title: Humanities II Name of The Lecturer: Mr. J.A.D.F.M. JayathilakaDocument8 pagesAssignment 2: Module Code: HN5001 Module Title: Humanities II Name of The Lecturer: Mr. J.A.D.F.M. JayathilakaDimuthu Nuwan AbeysingheNo ratings yet
- Extended Heredity: A New Understanding of Inheritance and EvolutionFrom EverandExtended Heredity: A New Understanding of Inheritance and EvolutionNo ratings yet
- The Evolution of Adaptive Systems: The General Theory of EvolutionFrom EverandThe Evolution of Adaptive Systems: The General Theory of EvolutionNo ratings yet
- Running Out Of Time: Introducing Behaviorology To Help Solve Global ProblemsFrom EverandRunning Out Of Time: Introducing Behaviorology To Help Solve Global ProblemsNo ratings yet
- Why (Almost All) Cosmologists Are AtheistsDocument14 pagesWhy (Almost All) Cosmologists Are Atheistsskd skdNo ratings yet
- Dickins & Dickins (Eds.) (2023) Evolutionary Biology - Contemporary and Historical Reflections Upon Core TheoryDocument606 pagesDickins & Dickins (Eds.) (2023) Evolutionary Biology - Contemporary and Historical Reflections Upon Core TheoryFabregasAlejandroNo ratings yet
- North American Philosophical PublicationsDocument25 pagesNorth American Philosophical PublicationsDon ArthurNo ratings yet
- Taylor & Francis, Ltd. British Journal of Sociology of EducationDocument21 pagesTaylor & Francis, Ltd. British Journal of Sociology of EducationSP DaniloNo ratings yet
- Evolutionary Biology and ReligionDocument23 pagesEvolutionary Biology and Religionpgkoutsoukos_7802726No ratings yet
- Odenbaugh&Griffith. Philosophy of BiologyDocument14 pagesOdenbaugh&Griffith. Philosophy of BiologyManu Rodríguez VázquezNo ratings yet
- Clayton, 1997, Philosophy of Science, What One Needs To Know PDFDocument10 pagesClayton, 1997, Philosophy of Science, What One Needs To Know PDFAna Chacón RosasNo ratings yet
- Calcott 2014Document12 pagesCalcott 2014raul roblesNo ratings yet
- Bergstrom 2009Document18 pagesBergstrom 2009raul roblesNo ratings yet
- Brandon 1997Document14 pagesBrandon 1997raul roblesNo ratings yet
- Bapteste 2012Document26 pagesBapteste 2012raul roblesNo ratings yet
- Baetu 2010Document18 pagesBaetu 2010raul roblesNo ratings yet
- The British Society For The Philosophy of Science, Oxford University Press The British Journal For The Philosophy of ScienceDocument18 pagesThe British Society For The Philosophy of Science, Oxford University Press The British Journal For The Philosophy of Scienceraul roblesNo ratings yet
- 10 2307@23334314Document13 pages10 2307@23334314raul roblesNo ratings yet
- The University of Chicago Press Philosophy of Science AssociationDocument20 pagesThe University of Chicago Press Philosophy of Science Associationraul roblesNo ratings yet
- Stazione Zoologica Anton Dohrn - NapoliDocument25 pagesStazione Zoologica Anton Dohrn - Napoliraul roblesNo ratings yet
- Biochem Eng ProblemsDocument23 pagesBiochem Eng ProblemsAdu GilbertNo ratings yet
- Pegda EnzymeDocument9 pagesPegda EnzymecarolasbdNo ratings yet
- Experiment 4.1 (1) BiologyDocument2 pagesExperiment 4.1 (1) BiologyFitri ZuhairiNo ratings yet
- Dahleh, 2021 PDFDocument13 pagesDahleh, 2021 PDFFani araujoNo ratings yet
- BookOfAbstracts 15thWORKSHOP PDFDocument392 pagesBookOfAbstracts 15thWORKSHOP PDFkillermonkeyNo ratings yet
- Specimen Mark Scheme Paper 4Document8 pagesSpecimen Mark Scheme Paper 4FatinAzahraNo ratings yet
- UntitledDocument8 pagesUntitledQudsia AbrarNo ratings yet
- Enzymes MCQDocument3 pagesEnzymes MCQSundaralingam Raj78% (9)
- Question 1 (37 Marks) : Biochemistry 3 BCH 314Document4 pagesQuestion 1 (37 Marks) : Biochemistry 3 BCH 314victorNo ratings yet
- Lecture On SensorsDocument106 pagesLecture On Sensorsapi-19959578No ratings yet
- Food Enzymology: FD-206 Spring Semester 2021 Second Year Food EngineeringDocument25 pagesFood Enzymology: FD-206 Spring Semester 2021 Second Year Food EngineeringSahar Batool QaziNo ratings yet
- Celina Nassar D: Biology Department Biology Worksheet 7 - Second Semester Name: SectionDocument37 pagesCelina Nassar D: Biology Department Biology Worksheet 7 - Second Semester Name: SectionCelina NassarNo ratings yet
- Bio Finish in TextilesDocument35 pagesBio Finish in Textilessahu_disha100% (2)
- Enzymes Mind MapDocument1 pageEnzymes Mind Mapruaridh.sayerNo ratings yet
- Manufacturing of Carrot JuiceDocument42 pagesManufacturing of Carrot JuiceKhoa Trần MinhNo ratings yet
- El Ejercicio Es Medicina, Pero ¿Interfiere Con La Medicina?Document9 pagesEl Ejercicio Es Medicina, Pero ¿Interfiere Con La Medicina?Agustin LopezNo ratings yet
- Ib Course PlannerDocument11 pagesIb Course PlannerTijana Tosic100% (1)
- Amit Kumar Sinha - Nutrizyme - Aquaculture HealthDocument40 pagesAmit Kumar Sinha - Nutrizyme - Aquaculture HealthMitshutopNo ratings yet
- 1 Microbial Metabolism andDocument103 pages1 Microbial Metabolism andcalocetcerphus509No ratings yet
- THC YeastDocument9 pagesTHC YeastAdrianMrózNo ratings yet
- Hsslive-XI-zoolgy-prvs Years Chapter Wise-Signed PDFDocument40 pagesHsslive-XI-zoolgy-prvs Years Chapter Wise-Signed PDFAkshayNo ratings yet
- Changes in Quality Attributes During Storage of High-Pressure and Thermally Processed Pineapple PureeDocument24 pagesChanges in Quality Attributes During Storage of High-Pressure and Thermally Processed Pineapple PureePaul Jefferson Flores HurtadoNo ratings yet
- Kuliah Pengantar Enzimologi Baru 1Document35 pagesKuliah Pengantar Enzimologi Baru 1okpitNo ratings yet
- Catalysis Notes PDFDocument12 pagesCatalysis Notes PDFbbtbadal100% (2)
- PapainDocument3 pagesPapainMohamed0% (1)
- Soil Enzymes PowerpointDocument71 pagesSoil Enzymes PowerpointAndrae Genus67% (6)
- Handling Toxicology Lab SamplesDocument6 pagesHandling Toxicology Lab SamplesEvans MunyaoNo ratings yet
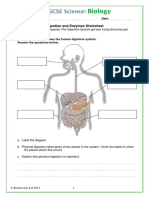
- Digestion and Enzymes Worksheet: 1. The Diagram Below Shows The Human Digestive System. Answer The Questions BelowDocument4 pagesDigestion and Enzymes Worksheet: 1. The Diagram Below Shows The Human Digestive System. Answer The Questions BelowHana Mohamed HanyNo ratings yet
- Experiment 4 (Biology)Document11 pagesExperiment 4 (Biology)言爱邦0% (1)
- Snape, Alison, Papachristodoulou, Despo, Elliott, William H., El - Biochemistry and Molecular Biology 6th Edition (2018, Oxford University Press) - Libgen - LiDocument641 pagesSnape, Alison, Papachristodoulou, Despo, Elliott, William H., El - Biochemistry and Molecular Biology 6th Edition (2018, Oxford University Press) - Libgen - LiĐinh Tiến ĐạtNo ratings yet