You might also like
- American Board of Orthodontics Flashcards - CramDocument112 pagesAmerican Board of Orthodontics Flashcards - Cramimtiyazh85100% (14)
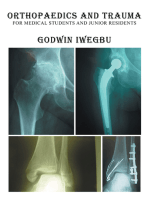
- Orthopaedics and Trauma for Medical Students and Junior ResidentsFrom EverandOrthopaedics and Trauma for Medical Students and Junior ResidentsRating: 5 out of 5 stars5/5 (3)
- Essentials of Skeletal Radiology - (Normal Skeletal Anatomy and Radiographic Positioning)Document196 pagesEssentials of Skeletal Radiology - (Normal Skeletal Anatomy and Radiographic Positioning)jdNo ratings yet
- Exsc 223 Practice Exam 2Document11 pagesExsc 223 Practice Exam 2kimber brownNo ratings yet
- Eizenberg's General AnatomyDocument255 pagesEizenberg's General Anatomymushroom620100% (9)
- Lecture Notes On Orthopaedics and Fractures PDFDocument257 pagesLecture Notes On Orthopaedics and Fractures PDFDanny Irung0% (1)
- Artículo Cuestionario 1Document10 pagesArtículo Cuestionario 1Carmen BarajasNo ratings yet
- Embryology of The Head and Neck: Martin CatalaDocument44 pagesEmbryology of The Head and Neck: Martin CatalaGita ArneniNo ratings yet
- 2017 Vascularised Bone TransferDocument11 pages2017 Vascularised Bone TransferRoger WatersNo ratings yet
- Collagens-Structure, Function, and Biosynthesis PDFDocument16 pagesCollagens-Structure, Function, and Biosynthesis PDFayudewintasariNo ratings yet
- Evolution of Molluscs 2019Document14 pagesEvolution of Molluscs 2019CoevicNo ratings yet
- Cunningham Et Al 2016 Chapter 3Document17 pagesCunningham Et Al 2016 Chapter 3Rodrigo Vivero MirandaNo ratings yet
- Alveolar Exostosis - Revisited A Narrative Review of The LiteratureDocument6 pagesAlveolar Exostosis - Revisited A Narrative Review of The Literatureغمدان دماج الحمزيNo ratings yet
- Liversidge 2016 in Dev Juv OsteolDocument29 pagesLiversidge 2016 in Dev Juv OsteolLidia IftimeNo ratings yet
- Biomechanical - Determinants - of - The - Stability of Dental ImplantsDocument11 pagesBiomechanical - Determinants - of - The - Stability of Dental ImplantsJuan Manuel Zegarra CuyaNo ratings yet
- Liversidge 2016 in Dev Juv OsteolDocument29 pagesLiversidge 2016 in Dev Juv OsteolflorenceliboyoNo ratings yet
- 1 Tooth - Development QuestionsDocument12 pages1 Tooth - Development QuestionsMena ZayanNo ratings yet
- Radaic 2021Document26 pagesRadaic 2021Elías Enrique MartínezNo ratings yet
- Yan JDocument111 pagesYan JDattatraya ParleNo ratings yet
- Alveolar Exostosis - Revisited: A Narrative Review of The LiteratureDocument6 pagesAlveolar Exostosis - Revisited: A Narrative Review of The LiteratureFransiski HoNo ratings yet
- Biological Basis of Distraction OsteogenesisDocument7 pagesBiological Basis of Distraction Osteogenesisdr.antonio.velasquezNo ratings yet
- Odontoblast PhysiologyDocument7 pagesOdontoblast PhysiologyWillian De Oliveira FrancaNo ratings yet
- FGHF Regenerative DentistryDocument178 pagesFGHF Regenerative DentistrybuzatugeorgescuNo ratings yet
- The Impact of Microgravity On Bone in HumansDocument13 pagesThe Impact of Microgravity On Bone in HumansVaibhav SinghNo ratings yet
- Anatomy Summary BasicDocument54 pagesAnatomy Summary BasicDungani AllanNo ratings yet
- A Brief Introduction Into Orthopaedic ImplantsDocument20 pagesA Brief Introduction Into Orthopaedic ImplantsLuisAngelPonceTorresNo ratings yet
- Osteocyte Function Under Compressive Mechanical Force: SciencedirectDocument11 pagesOsteocyte Function Under Compressive Mechanical Force: SciencedirectZachary DuongNo ratings yet
- Autoimmunity Reviews: J. Caetano-Lopes, H. Canhão, J.E. FonsecaDocument6 pagesAutoimmunity Reviews: J. Caetano-Lopes, H. Canhão, J.E. FonsecaNecróforo HernándezNo ratings yet
- Growth and Development 2Document10 pagesGrowth and Development 2Ali MONo ratings yet
- Journal of Oral Biosciences: Sumio NishikawaDocument8 pagesJournal of Oral Biosciences: Sumio NishikawaTatiana Benito RojasNo ratings yet
- Ingenieria Tejidos CorneaDocument29 pagesIngenieria Tejidos CorneaJose Luis Navia AvilaNo ratings yet
- Sella TurcicaDocument4 pagesSella TurcicaHeidar ZaarourNo ratings yet
- 1 s2.0 S0925477317308043 MainDocument10 pages1 s2.0 S0925477317308043 MainRogelio FloresNo ratings yet
- Ecoimmunology and Microbial Ecology Contributions To Avian BehaviorDocument10 pagesEcoimmunology and Microbial Ecology Contributions To Avian BehaviorLUZ ANDREA GARCIA CAYCEDONo ratings yet
- Soft Tissue Sealing Around Dental Implants Based On Histological InterpretationDocument9 pagesSoft Tissue Sealing Around Dental Implants Based On Histological Interpretationcmfvaldesr7No ratings yet
- Anatomy Department, Queen's University, BelfastDocument11 pagesAnatomy Department, Queen's University, BelfastEDWIN ARLEY MURILLO PEÑANo ratings yet
- The Stele - A Developmental Perspective On The Diversity and Evolution of Primary Vascular ArchitectureDocument27 pagesThe Stele - A Developmental Perspective On The Diversity and Evolution of Primary Vascular ArchitectureRickySaptarshi SahaNo ratings yet
- The Basic Principles of External Fixation Using The Ilizarov DeviceDocument30 pagesThe Basic Principles of External Fixation Using The Ilizarov DevicedendroaspisblackNo ratings yet
- Growth AssinmentDocument23 pagesGrowth Assinmentmohammed.eskanderNo ratings yet
- Femorals PDFDocument145 pagesFemorals PDFnoronisa talusobNo ratings yet
- Case Study On Open Tibial FractureDocument48 pagesCase Study On Open Tibial FractureOmotosho AlexNo ratings yet
- Osteochondral Tissue EngineeringDocument16 pagesOsteochondral Tissue EngineeringWookyoung LeeNo ratings yet
- Bedoya 2014Document10 pagesBedoya 2014Ortodoncia UnicocNo ratings yet
- Atm BiomecanicaDocument8 pagesAtm BiomecanicaJuan Carlos MeloNo ratings yet
- Biological Basis of Distraction Osteogenesis PDFDocument7 pagesBiological Basis of Distraction Osteogenesis PDFJuankNo ratings yet
- Solution Manual For Essentials of Anatomy and Physiology 1st by PattonDocument12 pagesSolution Manual For Essentials of Anatomy and Physiology 1st by PattonDeanBucktdjx100% (40)
- Biological Reviews - 2015 - Cellerino - From The Bush To The Bench The Annual Nothobranchius Fishes As A New Model SystemDocument23 pagesBiological Reviews - 2015 - Cellerino - From The Bush To The Bench The Annual Nothobranchius Fishes As A New Model SystemEmerson CasaliNo ratings yet
- Chapter 2: The Skeletal System - Osteology: Reading: Moore, Dalley, & Agur ObjectivesDocument10 pagesChapter 2: The Skeletal System - Osteology: Reading: Moore, Dalley, & Agur ObjectivesVarsha SNo ratings yet
- Livio Luzi (Auth.), Livio Luzi (Eds.) - Cellular Physiology and Metabolism of Physical Exercise-Springer-Verlag Mailand (2012)Document167 pagesLivio Luzi (Auth.), Livio Luzi (Eds.) - Cellular Physiology and Metabolism of Physical Exercise-Springer-Verlag Mailand (2012)Carol Estefany Cuevas PolancoNo ratings yet
- Procrsmed00410 0051 PDFDocument10 pagesProcrsmed00410 0051 PDFMilton David Rios SerratoNo ratings yet
- What Affects The Biocompatibility of PolymersDocument25 pagesWhat Affects The Biocompatibility of PolymersAndreaNo ratings yet
- A Brief Introduction Into Orthopaedic Implants: Screws, Plates, and NailsDocument21 pagesA Brief Introduction Into Orthopaedic Implants: Screws, Plates, and Nailsfebyan yohanesNo ratings yet
- Growth Site and Center - TextDocument9 pagesGrowth Site and Center - Textneven.mortagaNo ratings yet
- Maxillary Sinus Bone GraftingDocument181 pagesMaxillary Sinus Bone GraftingTuấn Hành TrầnNo ratings yet
- 1.16 (Surgery) Bone HealingDocument4 pages1.16 (Surgery) Bone HealingLeo Mari Go LimNo ratings yet
- Treatment of The Fracture of Femur in ChildrenDocument35 pagesTreatment of The Fracture of Femur in Childrenmoaz.zaiter.transporteNo ratings yet
- Ato ZOrthodontics Vol 2 GrowthanddevelopmentDocument17 pagesAto ZOrthodontics Vol 2 GrowthanddevelopmentambersNo ratings yet
- Anatomy and Biomechanics FinalDocument34 pagesAnatomy and Biomechanics FinalHasan Abo MohamedNo ratings yet
- Growth AssinmentDocument9 pagesGrowth Assinmentmohammed.eskanderNo ratings yet
- ImplantDocument46 pagesImplantMosica VladNo ratings yet
- Recent Advances in Heteromorph Ammonoid PalaeobiologyDocument35 pagesRecent Advances in Heteromorph Ammonoid PalaeobiologyJosé Juárez RuizNo ratings yet
- 1 s2.0 S1349007922000020 MainDocument11 pages1 s2.0 S1349007922000020 MainMuhammad GhufronNo ratings yet
- Bone Marrow MRI - A Pattern-Based Approach (PDFDrive)Document183 pagesBone Marrow MRI - A Pattern-Based Approach (PDFDrive)IrinaMariaStrugari100% (1)
- Maxilary Sinus Graft ArunDocument181 pagesMaxilary Sinus Graft ArunJose Fernando Fragozo MendozaNo ratings yet
- Advanced farriery knowledge: A study guide and AWCF theory course companionFrom EverandAdvanced farriery knowledge: A study guide and AWCF theory course companionNo ratings yet
- Physeal Closure Time & DiseaseDocument8 pagesPhyseal Closure Time & DiseaseMelissa StrawhunNo ratings yet
- Bone Physiology NotesDocument6 pagesBone Physiology NotesTAMMY CHARLENE HARTELNo ratings yet
- ChemistryDocument29 pagesChemistryIshkruti WaghmareNo ratings yet
- Anatomy and Healing Process of Paediatric Bone - Dato DR Zulkiflee OsmanDocument56 pagesAnatomy and Healing Process of Paediatric Bone - Dato DR Zulkiflee OsmanMuhammad Naqiuddin JalaluddinNo ratings yet
- Chapter 6 Skeletal System 1Document18 pagesChapter 6 Skeletal System 1M GarciaNo ratings yet
- HANDOUTS Prelim CH 6Document40 pagesHANDOUTS Prelim CH 6Abia Annieson A. LorenzoNo ratings yet
- Bones and Bone TissueDocument12 pagesBones and Bone TissueJharaNo ratings yet
- 1basic Concepts of Growth and DevelopmentDocument100 pages1basic Concepts of Growth and Developmentshweta nageshNo ratings yet
- Anatomy: Petr Hájek, MD, Phd. Head of Anatomy Dept. Assoc. Prof. Dáša Slížová, MD, Phd. Senior LecturerDocument27 pagesAnatomy: Petr Hájek, MD, Phd. Head of Anatomy Dept. Assoc. Prof. Dáša Slížová, MD, Phd. Senior LecturerTodesengelNo ratings yet
- Bone Healing, Regeneration and Repair: On BiomolecoulerDocument26 pagesBone Healing, Regeneration and Repair: On BiomolecoulerSarah Nabella PutriNo ratings yet
- Connective TissueDocument202 pagesConnective TissuePeter ThompsonNo ratings yet
- Lecture: Bone DepositionDocument19 pagesLecture: Bone Depositionapi-3798860No ratings yet
- Fracture !!!!!!Document31 pagesFracture !!!!!!Maryjoy MertallaNo ratings yet
- Embryology 6 Limb DevelopmentDocument17 pagesEmbryology 6 Limb DevelopmentVokeh Dante MedDeeNo ratings yet
- Human Anatomy 5Th Edition Saladin Solutions Manual Full Chapter PDFDocument28 pagesHuman Anatomy 5Th Edition Saladin Solutions Manual Full Chapter PDFJenniferWhitebctr100% (11)
- Chemistry Project On Estimation of Contant of Bone Ash PDFDocument9 pagesChemistry Project On Estimation of Contant of Bone Ash PDFPooja BhagwatNo ratings yet
- Alveolar Bone in Health and DiseaseDocument16 pagesAlveolar Bone in Health and DiseaseVartika VermaNo ratings yet
- Anatomy Skeletal Tissue - Chap 5Document30 pagesAnatomy Skeletal Tissue - Chap 5pancit cantonNo ratings yet
- Low Intensity Pulsed Ultrasound 2012Document12 pagesLow Intensity Pulsed Ultrasound 2012Joseph SmithNo ratings yet
- Case Study Osteo YnbbDocument11 pagesCase Study Osteo YnbbAyunie ZulkeflyNo ratings yet
- Compilation AnaphyDocument219 pagesCompilation AnaphyIsabel Bibat DavidNo ratings yet
- Classification of Bones On The Basis of ShapeDocument9 pagesClassification of Bones On The Basis of ShapeR Hornilla ArcegaNo ratings yet
- Biology Chapter 13 Concepts 01 35 Human Biology Answers PDFDocument145 pagesBiology Chapter 13 Concepts 01 35 Human Biology Answers PDFKamryn MontgomeryNo ratings yet
- Genanat D&R AgamDocument51 pagesGenanat D&R AgamWitty GreenNo ratings yet
- AO Surgery ReferenceDocument8 pagesAO Surgery ReferenceHurulaeniNo ratings yet