You might also like
- The Hippocampus BookDocument853 pagesThe Hippocampus BookLucia Lopez100% (3)
- The Limbic Brain. Andrew Lautin PDFDocument152 pagesThe Limbic Brain. Andrew Lautin PDFAntonio D LazodelaVegaNo ratings yet
- Limbic Sys - ArchDocument85 pagesLimbic Sys - Archdrkadiyala2No ratings yet
- Clinical: Dialogu S eDocument144 pagesClinical: Dialogu S eDiego A. Cuadros Torres100% (3)
- Squire Zola 1996Document8 pagesSquire Zola 1996cecilia martinezNo ratings yet
- Kovner, Et Al - Cortico-Limbic Interactions Relevant To Psychopathology (2019)Document13 pagesKovner, Et Al - Cortico-Limbic Interactions Relevant To Psychopathology (2019)Ricardo EscNo ratings yet
- AMM 64 S1 WebDocument176 pagesAMM 64 S1 WebAndrada CatrinoiuNo ratings yet
- 1 s2.0 S096999612030406X MainDocument12 pages1 s2.0 S096999612030406X MainLuis ricardo Véjar felixNo ratings yet
- Spike Protein Makes Blood Brain Barrier Mopre Porus, PaperDocument13 pagesSpike Protein Makes Blood Brain Barrier Mopre Porus, PaperMareseatoatsNo ratings yet
- Neurology and Covid 2Document20 pagesNeurology and Covid 2Novi SeptianiNo ratings yet
- Neuro-COVID Is Not at Variance Between Children and AdultsDocument3 pagesNeuro-COVID Is Not at Variance Between Children and AdultsMaurycy RakowskiNo ratings yet
- Variation of Cognition and Cerebral Activity With Severity of Airflow Obstruction in CopdDocument9 pagesVariation of Cognition and Cerebral Activity With Severity of Airflow Obstruction in CopdIJAR JOURNALNo ratings yet
- A Comprehensive Review of Neurologic Manifestations of COVID-19 and Management of Pre-Existing Neurologic Disorders in ChildrenDocument7 pagesA Comprehensive Review of Neurologic Manifestations of COVID-19 and Management of Pre-Existing Neurologic Disorders in ChildrenNoviaNo ratings yet
- (2020-JAMA) - Neuropathogenesis and Neurologic Manifestations of The Coronaviruses in The Age of Coronavirus Disease 2019Document10 pages(2020-JAMA) - Neuropathogenesis and Neurologic Manifestations of The Coronaviruses in The Age of Coronavirus Disease 2019Iván PérezNo ratings yet
- Near Infrared Spectroscopy For Assessing OxygenDocument6 pagesNear Infrared Spectroscopy For Assessing OxygenInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Artigo 5Document5 pagesArtigo 5Paloma PaixãoNo ratings yet
- Pereira MSDocument10 pagesPereira MSAntónio AlvimNo ratings yet
- Ebiomedicine: Qi Cheng, Yue Yang, Jianqun GaoDocument6 pagesEbiomedicine: Qi Cheng, Yue Yang, Jianqun Gaowedib17801No ratings yet
- SARS CoV 2 Brain StructureDocument26 pagesSARS CoV 2 Brain StructureJoseNo ratings yet
- Basal Ganglia Involvement and AlteredDocument8 pagesBasal Ganglia Involvement and AlteredJonnathan Quintero (tatán)No ratings yet
- A Review of Neurological Involvement in Patients With Sars-Cov-2 InfectionDocument11 pagesA Review of Neurological Involvement in Patients With Sars-Cov-2 Infectionakjsndka aksdkasndNo ratings yet
- Pathophysiology-And-Management-Of-Neurogenic-Pulmonary-Edema-In-Patients-With-Acute-Severe-Brain-Injury EspañolDocument7 pagesPathophysiology-And-Management-Of-Neurogenic-Pulmonary-Edema-In-Patients-With-Acute-Severe-Brain-Injury EspañolOscar SanNo ratings yet
- Alzheimers Selkoe Science 2012 PDFDocument5 pagesAlzheimers Selkoe Science 2012 PDFLong TomNo ratings yet
- Articulo CovidDocument14 pagesArticulo Coviddanyescalerapumas24No ratings yet
- Covid 19Document8 pagesCovid 19Bhagass BhagassNo ratings yet
- As Sequelas Neuropsiquiátricas Crônicas de COVID-19 - Possíveis Impactos em Demências e DADocument10 pagesAs Sequelas Neuropsiquiátricas Crônicas de COVID-19 - Possíveis Impactos em Demências e DAAlberto HenriqueNo ratings yet
- CNS 27 36Document12 pagesCNS 27 36Isabely AmabilyNo ratings yet
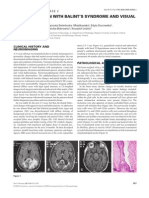
- 31-Year-Old Man With Balint'S Syndrome and Visual Problems: Clinical History and NeuroimagingDocument4 pages31-Year-Old Man With Balint'S Syndrome and Visual Problems: Clinical History and NeuroimagingwillygopeNo ratings yet
- Biomedicines 11 00044 v3Document19 pagesBiomedicines 11 00044 v3mariel gonzalezNo ratings yet
- Nanomedicine: A Promising Way To Manage Alzheimer's DiseaseDocument20 pagesNanomedicine: A Promising Way To Manage Alzheimer's Diseasesara salamaNo ratings yet
- Physiology & Behavior: Tong Wang, Cun-Zhi Liu, Jian-Chun Yu, Wen Jiang, Jing-Xian HanDocument7 pagesPhysiology & Behavior: Tong Wang, Cun-Zhi Liu, Jian-Chun Yu, Wen Jiang, Jing-Xian Handouglasvargas-1No ratings yet
- Bell 'S Palsy: Clinical and Neurophysiologic Predictors of RecoveryDocument5 pagesBell 'S Palsy: Clinical and Neurophysiologic Predictors of RecoveryMarsya Yulinesia LoppiesNo ratings yet
- Endocannabinoid System Unlocks The Puzzle of Autism Treatment Via MicrogliaDocument10 pagesEndocannabinoid System Unlocks The Puzzle of Autism Treatment Via MicrogliaMicaela SaNchezNo ratings yet
- 10 - Elucidating The Neuropathologic Mechanisms of SARS-CoV-2 InfectionDocument19 pages10 - Elucidating The Neuropathologic Mechanisms of SARS-CoV-2 InfectionVerónica GaticaNo ratings yet
- Chronic SubduralDocument6 pagesChronic Subduralshifa tropmedNo ratings yet
- Asian Journal of PsychiatryDocument4 pagesAsian Journal of PsychiatryCogvid ProjectNo ratings yet
- Ischemic Cerebral Stroke Case Report, Complications and Associated FactorsDocument7 pagesIschemic Cerebral Stroke Case Report, Complications and Associated FactorsAlfa DexterNo ratings yet
- Cerebral Autoregulation Indices Are Not Interchangeable in Patients With SepsisDocument7 pagesCerebral Autoregulation Indices Are Not Interchangeable in Patients With SepsisFrank Antony Arizabal GaldosNo ratings yet
- Angiotensin-Converting Enzyme 2 (ACE2) Expression Increases With Age in Patients Requiring Mechanical VentilationDocument17 pagesAngiotensin-Converting Enzyme 2 (ACE2) Expression Increases With Age in Patients Requiring Mechanical VentilationJuan Vera SanchezNo ratings yet
- Daniel PiriciDocument12 pagesDaniel PiriciRashellya RasyidaNo ratings yet
- Complicaciones Neurológicas de La COVID-19 Desde La Fisiopatología Hasta La Rehabilitación. Una Visión GeneralDocument17 pagesComplicaciones Neurológicas de La COVID-19 Desde La Fisiopatología Hasta La Rehabilitación. Una Visión GeneralB10. Oscar Alonso Díaz PintoNo ratings yet
- Journal AlzheimerDocument10 pagesJournal AlzheimerFaza KeumalasariNo ratings yet
- Neuroscience Letters: John Kealy, Chris Greene, Matthew Campbell TDocument14 pagesNeuroscience Letters: John Kealy, Chris Greene, Matthew Campbell TAndré AlexandreNo ratings yet
- Sars-Cov2 Spike Protein Accumulation in The Skull-Meninges Brain Axis PDFDocument26 pagesSars-Cov2 Spike Protein Accumulation in The Skull-Meninges Brain Axis PDFemmanuel AndemNo ratings yet
- Graphical SummaryDocument26 pagesGraphical Summaryemmanuel AndemNo ratings yet
- Origins of Cerebral Edema: Implications For Space Ight-Associated Neuro-Ocular SyndromeDocument8 pagesOrigins of Cerebral Edema: Implications For Space Ight-Associated Neuro-Ocular SyndromeDaniel PadillaNo ratings yet
- Main - CópiaDocument7 pagesMain - CópiaMariana OliveiraNo ratings yet
- Fnagi 14 800617Document10 pagesFnagi 14 800617Snezana MihajlovicNo ratings yet
- Role of Endothelial Cell Receptors in The Context of SARS CoV 2 Infection COVID 19Document8 pagesRole of Endothelial Cell Receptors in The Context of SARS CoV 2 Infection COVID 19Doménica SánchezNo ratings yet
- 14 Brain and Behavior - 2023 - Li - Causal Effect of Psychiatric Disorders On Epilepsy A Two Sample Mendelian RandomizationDocument9 pages14 Brain and Behavior - 2023 - Li - Causal Effect of Psychiatric Disorders On Epilepsy A Two Sample Mendelian RandomizationDAFNE FRANÇA SANTANANo ratings yet
- Long-COVID - Cognitive Deficits (Brain Fog) and Brain Lesions in Non-Hospitalized Patients - PMCDocument4 pagesLong-COVID - Cognitive Deficits (Brain Fog) and Brain Lesions in Non-Hospitalized Patients - PMCMaria SilvaNo ratings yet
- Biomarkers For Alzheimer's DiseaseDocument13 pagesBiomarkers For Alzheimer's DiseaseFernando lazzarettiNo ratings yet
- JURNALDocument10 pagesJURNALLuses Shantia HaryantoNo ratings yet
- Blood-Brain Barrier Disruption and Sustained Systemic Inflammation in Individuals With Long COVID-associated Cognitive ImpairmentDocument27 pagesBlood-Brain Barrier Disruption and Sustained Systemic Inflammation in Individuals With Long COVID-associated Cognitive ImpairmentKAMI LEGENDNo ratings yet
- Post-Viral Effects of COVID-19 in The Olfactory System and Their ImplicationsDocument9 pagesPost-Viral Effects of COVID-19 in The Olfactory System and Their Implicationschan kyoNo ratings yet
- COVID-19 Sleep, Circadian Rhythms and ImmunityDocument22 pagesCOVID-19 Sleep, Circadian Rhythms and ImmunityFrancisco MtzNo ratings yet
- Fneur 11 00071Document10 pagesFneur 11 00071Nurvia AndrianiNo ratings yet
- 2-A Comprehensive Investigation of The MRNA and Protein Level of ACE2, The Putative Receptor of SARS-CoV-2, in Human Tissues and Blood CellsDocument10 pages2-A Comprehensive Investigation of The MRNA and Protein Level of ACE2, The Putative Receptor of SARS-CoV-2, in Human Tissues and Blood CellschimansqmNo ratings yet
- EBJ ArticleDocument7 pagesEBJ ArticleKhristle DavidNo ratings yet
- BloodBrainBarrier LongCovid s41593 024 01576 9Document27 pagesBloodBrainBarrier LongCovid s41593 024 01576 9gjlynxNo ratings yet
- Inflammatory Arthritis and Younger Age of Lupus Onset Are Associated With The Development of Neuropsychiatric LupusDocument5 pagesInflammatory Arthritis and Younger Age of Lupus Onset Are Associated With The Development of Neuropsychiatric LupusHas SimNo ratings yet
- 2018 Article 1097Document4 pages2018 Article 1097rajaririnNo ratings yet
- Mixed Central and Peripheral Nervous System Disorders in Severe Sars Cov 2 InfectionDocument7 pagesMixed Central and Peripheral Nervous System Disorders in Severe Sars Cov 2 Infectionchartreuse avonleaNo ratings yet
- 932 Full-2Document10 pages932 Full-2sara laadamiNo ratings yet
- MiscDocument8 pagesMiscTajul TajNo ratings yet
- Badaut Zhang Preface2019 ADocument5 pagesBadaut Zhang Preface2019 AGonzalo AltamiranoNo ratings yet
- Clinical Neuroimmunology: Multiple Sclerosis and Related DisordersFrom EverandClinical Neuroimmunology: Multiple Sclerosis and Related DisordersNo ratings yet
- Physiology 5 - EpilepsyDocument17 pagesPhysiology 5 - EpilepsyTomgfgd RogestoreNo ratings yet
- Anterior Posterior HippocampusDocument25 pagesAnterior Posterior HippocampusChrysoula GkaniNo ratings yet
- Giuliano Et Al-2017-Hippocampus PDFDocument14 pagesGiuliano Et Al-2017-Hippocampus PDFdess101No ratings yet
- Section Properties Torsional Constant Area of SectionDocument38 pagesSection Properties Torsional Constant Area of SectionAlma M. LaraNo ratings yet
- Schema Ketronx1Document12 pagesSchema Ketronx1Manrolla89% (9)
- Article PDFDocument50 pagesArticle PDFEricWallachNo ratings yet
- Macroanatomy and Microanatomy of The Temporal Lobe 2007Document12 pagesMacroanatomy and Microanatomy of The Temporal Lobe 2007Juan José Pabón CastroNo ratings yet
- Interaural 15.70 MM Bregma 6.70 MMDocument79 pagesInteraural 15.70 MM Bregma 6.70 MMSergey A. SavelyevNo ratings yet
- BE Ist Year Sem 1 and 2 - Full Scheme With Syllabus & SU CodeDocument86 pagesBE Ist Year Sem 1 and 2 - Full Scheme With Syllabus & SU CodePÀRMAR parthNo ratings yet
- Paxinos The Rat Brain 73Document1 pagePaxinos The Rat Brain 73joseuri2011No ratings yet
- Alekls 1202Document113 pagesAlekls 1202Joseph SikatumbaNo ratings yet
- Reynato Wijaya Saputro, Sunarno, Siti Muflichatun MardiatiDocument14 pagesReynato Wijaya Saputro, Sunarno, Siti Muflichatun MardiatiKholisotur RosyidahNo ratings yet
- Unbiased Stereological Estimation of The Number Neurons The Human HippocampusDocument22 pagesUnbiased Stereological Estimation of The Number Neurons The Human HippocampusRodrigo Felipe Toro MellaNo ratings yet
- Nguyễn Thành Cao-20150310 - BTVNDocument6 pagesNguyễn Thành Cao-20150310 - BTVNOnePiece FanNo ratings yet
- Striedter - 2015 - Evolution of The Hippocampus in Reptiles and BirdsDocument22 pagesStriedter - 2015 - Evolution of The Hippocampus in Reptiles and BirdsOsny SillasNo ratings yet
- Brain Regions With Definitions - NeuroLexDocument32 pagesBrain Regions With Definitions - NeuroLexSrinivas RajanalaNo ratings yet
- 1 s2.0 S0014488617300018 MainDocument11 pages1 s2.0 S0014488617300018 MainJingyun WuNo ratings yet
- Sci & Tech (Till Jan 2016)Document270 pagesSci & Tech (Till Jan 2016)Diptendu DasNo ratings yet
- 1 s2.0 S0891061819301024 MainDocument10 pages1 s2.0 S0891061819301024 MainRenato Guerra GonzalezNo ratings yet
- Paraphilias As A Sub Type of Obsessive-Compulsive Disorder: A Hypothetical Bio-Social ModelDocument14 pagesParaphilias As A Sub Type of Obsessive-Compulsive Disorder: A Hypothetical Bio-Social ModelmuiNo ratings yet
- Sherman Et Al 2023 Multiple Memory Subsystems Reconsidering Memory in The Mind and BrainDocument23 pagesSherman Et Al 2023 Multiple Memory Subsystems Reconsidering Memory in The Mind and BrainChris InNottsNo ratings yet
- DM Neurology ThesisDocument7 pagesDM Neurology ThesisFindSomeoneToWriteMyPaperLowell100% (2)