You might also like
- Grade 10 Terms and DefinitionsDocument11 pagesGrade 10 Terms and Definitionskhotso100% (6)
- Hydrocarbons Chapter 13 ReviewDocument70 pagesHydrocarbons Chapter 13 ReviewGururaj Vasisth100% (3)
- Gamsat Chemistry Sample QuestionsDocument6 pagesGamsat Chemistry Sample QuestionsM S Rahman100% (1)
- Earth and Science PointersDocument3 pagesEarth and Science PointersDominico Jr Pojas TanNo ratings yet
- General Piping Design: Document No. Rev.: A BMR-PIP-DBS-ST-0001 Page 1 of 20Document20 pagesGeneral Piping Design: Document No. Rev.: A BMR-PIP-DBS-ST-0001 Page 1 of 20Agus SupriadiNo ratings yet
- Production of Animal Feed Concentrates From Sour Sop and Pineapple Peels Using Solid State Fermentation.Document5 pagesProduction of Animal Feed Concentrates From Sour Sop and Pineapple Peels Using Solid State Fermentation.Gregory Kalona100% (1)
- Starter Cultures in Food ProductionFrom EverandStarter Cultures in Food ProductionBarbara SperanzaNo ratings yet
- FES Aspergillus 5Document9 pagesFES Aspergillus 5Alexis RodriguezNo ratings yet
- Paper 16Document6 pagesPaper 16Sorin LazarNo ratings yet
- Production of Amylase Enzyme From Mangrove Fungal Isolates: African Journal of BiotechnologyDocument9 pagesProduction of Amylase Enzyme From Mangrove Fungal Isolates: African Journal of BiotechnologyAliyah SajaNo ratings yet
- BioRes 08-1-0250 Hariharan N Opt Lignin Peroxidase SSF Pineapple Leaf 3060Document22 pagesBioRes 08-1-0250 Hariharan N Opt Lignin Peroxidase SSF Pineapple Leaf 3060Hildan H GNo ratings yet
- 2891 6404 1 PBDocument8 pages2891 6404 1 PBIsworo RukmiNo ratings yet
- Seed starches as pharmaceutical sourcesDocument6 pagesSeed starches as pharmaceutical sourcesChem EngNo ratings yet
- Journal Type (Thesis)Document12 pagesJournal Type (Thesis)api-19649313No ratings yet
- Supported By: Sample Collection and PreparationDocument5 pagesSupported By: Sample Collection and Preparationmoses richardNo ratings yet
- Cellulolytic Fruits Wastes: A Potential Support For Enzyme Assisted Protein ProductionDocument9 pagesCellulolytic Fruits Wastes: A Potential Support For Enzyme Assisted Protein ProductionkesavarajNo ratings yet
- Jurnal Acara 1Document6 pagesJurnal Acara 1dinaserevinaNo ratings yet
- Journal Type (Thesis)Document12 pagesJournal Type (Thesis)api-19649313No ratings yet
- Aspergillus FlavusDocument6 pagesAspergillus FlavusFebri ShaarawyNo ratings yet
- Isolation, Identification and Characterisation of The Dominant Microorganisms of Kule Naoto: The Maasai Traditional Fermented Milk in KenyaDocument10 pagesIsolation, Identification and Characterisation of The Dominant Microorganisms of Kule Naoto: The Maasai Traditional Fermented Milk in KenyaVivek MokanNo ratings yet
- Ajol File Journals - 413 - Articles - 203082 - Submission - Proof - 203082 4921 508128 1 10 20210126Document12 pagesAjol File Journals - 413 - Articles - 203082 - Submission - Proof - 203082 4921 508128 1 10 20210126iqra sadiqNo ratings yet
- B9387CC3447Document5 pagesB9387CC3447cemz5No ratings yet
- MAB - 2018 A PDFDocument6 pagesMAB - 2018 A PDFAmriNo ratings yet
- ISFAS 2012-SukirahDocument4 pagesISFAS 2012-SukirahSukirah Abdul RahmanNo ratings yet
- Biodegradation of Agricultural Wastes (Rice Straw and Sorghum Stalk) Into Substrates of Utilizable Products Using White Rot FungusDocument7 pagesBiodegradation of Agricultural Wastes (Rice Straw and Sorghum Stalk) Into Substrates of Utilizable Products Using White Rot FungusInes HrastinskiNo ratings yet
- Isolation and Characterization of Starter Culture From Spontaneous Fermentation of SourdoughDocument4 pagesIsolation and Characterization of Starter Culture From Spontaneous Fermentation of SourdoughHusna AdilaNo ratings yet
- Article1380180775 - Oyeleke and OduwoleDocument4 pagesArticle1380180775 - Oyeleke and Oduwoleaditi_joshee419No ratings yet
- Solid State Fermentation of Aspergillus Oryzae For Glucoamylase Production On Agro ResiduesDocument10 pagesSolid State Fermentation of Aspergillus Oryzae For Glucoamylase Production On Agro ResiduesArlunandaAdhiarthaNo ratings yet
- Research ArticleDocument4 pagesResearch ArticleMusfeera KhanNo ratings yet
- Amylase and Xylanase Production by Soil FungiDocument3 pagesAmylase and Xylanase Production by Soil FungiSubhadra JhaNo ratings yet
- Banana Waste As Substrate For A-Amylase Production by Bacillus Subtilis (CBTK 106) Under Solid-State FermentationDocument6 pagesBanana Waste As Substrate For A-Amylase Production by Bacillus Subtilis (CBTK 106) Under Solid-State FermentationAlejandro Murillo V.No ratings yet
- Bacillus Licheniformis BT5.9 Isolated From Changar Hot SpringDocument14 pagesBacillus Licheniformis BT5.9 Isolated From Changar Hot SpringAleivi PérezNo ratings yet
- Aspergillus Oryzae Extract Improve Bromegrass Degradation With Rumen BateriaDocument6 pagesAspergillus Oryzae Extract Improve Bromegrass Degradation With Rumen BateriaAndres BelalcazarNo ratings yet
- Growth Response of Activities ofDocument6 pagesGrowth Response of Activities ofSbenmohamedNo ratings yet
- Measurement of β-glucan and other nutritional characteristics in distinct strains of AgaricusDocument7 pagesMeasurement of β-glucan and other nutritional characteristics in distinct strains of AgaricusDaniela ChalarcaNo ratings yet
- El Uso de Pectinasa para Tratamiendo de Jugo de Manzana y Mora AzulDocument7 pagesEl Uso de Pectinasa para Tratamiendo de Jugo de Manzana y Mora AzulLaura RDNo ratings yet
- Optimized Solid State Fermentation Medium Enhances The Multienzymes Production From Penicillium Citrinum and AspergillusDocument15 pagesOptimized Solid State Fermentation Medium Enhances The Multienzymes Production From Penicillium Citrinum and AspergillusSerafín Pérez ContrerasNo ratings yet
- Production and application of amylases from Rhizopus for glucose syrupDocument9 pagesProduction and application of amylases from Rhizopus for glucose syrupbarbarahNo ratings yet
- Digestive Amylase Activity in Wheat Bug StagesDocument8 pagesDigestive Amylase Activity in Wheat Bug Stagespharmacologist786No ratings yet
- Stenochlaena Palustri 02Document8 pagesStenochlaena Palustri 02donny sophandiNo ratings yet
- 128925-Article Text-349303-1-10-20160120Document9 pages128925-Article Text-349303-1-10-20160120Richard Ivan Medina HoyosNo ratings yet
- Chenodium QuinoaDocument8 pagesChenodium QuinoaЛуизАпазаТ.No ratings yet
- Sci - HubDocument7 pagesSci - HubVõ Hữu Hồng ChiNo ratings yet
- Application of A Statistical DesignDocument8 pagesApplication of A Statistical DesignTheophile IZERENo ratings yet
- Comparative Prod of Amylase Enzyme From RhizopusDocument4 pagesComparative Prod of Amylase Enzyme From RhizopusIsworo RukmiNo ratings yet
- Chemical Composition, Physicochemical and Functional Properties of Custard Apple (Annona Squmosa) Seed Flours and Protein IsolateDocument5 pagesChemical Composition, Physicochemical and Functional Properties of Custard Apple (Annona Squmosa) Seed Flours and Protein IsolateInnovative Research PublicationsNo ratings yet
- Honey CropDocument6 pagesHoney CropashatjhangNo ratings yet
- Salawu 2011Document7 pagesSalawu 2011Eti ApriyantiNo ratings yet
- Pectin From Ripe Peels of Mango CultivarsDocument10 pagesPectin From Ripe Peels of Mango CultivarsIsabel Tacunan CastilloNo ratings yet
- Nicanor 2005 PDFDocument9 pagesNicanor 2005 PDFLorenaNo ratings yet
- Biradar Et Al 2016 - BiologiaDocument9 pagesBiradar Et Al 2016 - BiologiaRahul KhandareNo ratings yet
- Production and Optimization of Xylanase and α-Amylase from Non-Saccharomyces Yeasts (Pichia membranifaciens)Document10 pagesProduction and Optimization of Xylanase and α-Amylase from Non-Saccharomyces Yeasts (Pichia membranifaciens)Jefri Nur HidayatNo ratings yet
- Production and Optimization of Xylanase and α-Amylase from Non-Saccharomyces Yeasts (Pichia membranifaciens)Document10 pagesProduction and Optimization of Xylanase and α-Amylase from Non-Saccharomyces Yeasts (Pichia membranifaciens)Jefri Nur HidayatNo ratings yet
- Canaliculata (Ampullariidae) : Oxicity of Barringtonia Racemosa (L.) Kernel Extract On PomaceaDocument5 pagesCanaliculata (Ampullariidae) : Oxicity of Barringtonia Racemosa (L.) Kernel Extract On PomaceaDarmansyah YasinNo ratings yet
- Phytochemical Analysis and Cytotoxicity Studies of Curcuma Amada Rhizomes in BHK-21 CellsDocument7 pagesPhytochemical Analysis and Cytotoxicity Studies of Curcuma Amada Rhizomes in BHK-21 CellsJessica ClarkNo ratings yet
- Via Suspension Cultures of Banana (Musa SPP.) : Highly Efficient Somatic Embryogenesis and Plant RegenerationDocument12 pagesVia Suspension Cultures of Banana (Musa SPP.) : Highly Efficient Somatic Embryogenesis and Plant Regenerationth_kiranNo ratings yet
- Decolourization of Azo Dye Methyl Red byDocument7 pagesDecolourization of Azo Dye Methyl Red byEvelyn NathaliaNo ratings yet
- Amylase Activity of Starch Degrading BacDocument13 pagesAmylase Activity of Starch Degrading BacRaisya RagilNo ratings yet
- RS - Volume 9 - Issue 4 - Pages 378-391Document14 pagesRS - Volume 9 - Issue 4 - Pages 378-391samirNo ratings yet
- Optimization of Culture Condition in The Production of Bioenzymes by Bacteria Isolated From Poultry Waste in Sokoto State, NigeriaDocument8 pagesOptimization of Culture Condition in The Production of Bioenzymes by Bacteria Isolated From Poultry Waste in Sokoto State, NigeriaUMYU Journal of Microbiology Research (UJMR)No ratings yet
- Bio Tech - IJBTR - Determination of Tropane Alkaloids in - Ayman Amin - EgyptDocument10 pagesBio Tech - IJBTR - Determination of Tropane Alkaloids in - Ayman Amin - EgyptTJPRC PublicationsNo ratings yet
- Abstract 7 12Document6 pagesAbstract 7 12Nikita RizkyNo ratings yet
- 225 2017-CJFSDocument8 pages225 2017-CJFSYuliet SusantoNo ratings yet
- In Vitro Propagation of Stevia rebaudianaDocument11 pagesIn Vitro Propagation of Stevia rebaudianahenry72No ratings yet
- Modern Alkaloids: Structure, Isolation, Synthesis, and BiologyFrom EverandModern Alkaloids: Structure, Isolation, Synthesis, and BiologyErnesto FattorussoNo ratings yet
- Test For Hydrocarbons: Laboratory Activity CHEM 525Document39 pagesTest For Hydrocarbons: Laboratory Activity CHEM 525Jherby TeodoroNo ratings yet
- Zinc Sulfate Oral SolutionDocument1 pageZinc Sulfate Oral SolutionKasidit SornchaiNo ratings yet
- THIRD YEAR QUESTION 2015 Calcutta UniversityDocument4 pagesTHIRD YEAR QUESTION 2015 Calcutta UniversityUshnish RoyNo ratings yet
- HPLC Columns For Carbohydrates: Product SpecificationDocument2 pagesHPLC Columns For Carbohydrates: Product SpecificationSandip BasuNo ratings yet
- Earth Science Lecture 2 V.2Document39 pagesEarth Science Lecture 2 V.2MieNo ratings yet
- Acidity in TeaDocument17 pagesAcidity in TeaRukmani 1011No ratings yet
- Halogens (Group VIIA) PresentationDocument12 pagesHalogens (Group VIIA) PresentationPatrick RegidorNo ratings yet
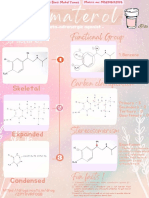
- Cimaterol Functional Groups and Carbon ClassificationDocument1 pageCimaterol Functional Groups and Carbon ClassificationHanny Mohd Yunus100% (1)
- PWD S.R. 2016-17 - KalaburagiDocument429 pagesPWD S.R. 2016-17 - KalaburagiKiran Rathod RitesNo ratings yet
- Preservation and Conservation TechniquesDocument20 pagesPreservation and Conservation TechniquesNur AthifahNo ratings yet
- Determining The Susceptibility of Stainless Steels and Related Nickel-Chromium-Iron Alloys To Stress-Corrosion Cracking in Polythionic AcidsDocument3 pagesDetermining The Susceptibility of Stainless Steels and Related Nickel-Chromium-Iron Alloys To Stress-Corrosion Cracking in Polythionic AcidsClaudio Di GioseffoNo ratings yet
- Industrial Training Performed at Piramal Pharma Solutions, Mahad, Dist-RaigadDocument32 pagesIndustrial Training Performed at Piramal Pharma Solutions, Mahad, Dist-RaigadChandan A. Wagh100% (1)
- Practical Organic Chemistry Classification TestsDocument19 pagesPractical Organic Chemistry Classification TestsJonathan ParkerNo ratings yet
- EU4 D2 M4 Inspection in Composites PDFDocument20 pagesEU4 D2 M4 Inspection in Composites PDFaerorenNo ratings yet
- The Punjab School Teaching Material of Class VIII Chapter 1 Human Organ Systems Q.no.1Document12 pagesThe Punjab School Teaching Material of Class VIII Chapter 1 Human Organ Systems Q.no.1Qulb e AbbasNo ratings yet
- CBSE Class 10 Science MCQ Bank Term IDocument48 pagesCBSE Class 10 Science MCQ Bank Term IAtharva Bhokare100% (1)
- Reactions of Carboxylic Acids and Its DerivativesDocument40 pagesReactions of Carboxylic Acids and Its DerivativesRoger ReyesNo ratings yet
- Utilization of Ecofriendly Self-Cleaning Concrete Using Zinc Oxide and Polypropylene FibreDocument3 pagesUtilization of Ecofriendly Self-Cleaning Concrete Using Zinc Oxide and Polypropylene FibreUmesha KempegowdaNo ratings yet
- MPHARM QA 1Y 2S 202T Pharm - ValidationDocument562 pagesMPHARM QA 1Y 2S 202T Pharm - Validationhosein bagheriNo ratings yet
- Life Cycle Assessment of Fertilizer Manufacturing and Impacts (39Document31 pagesLife Cycle Assessment of Fertilizer Manufacturing and Impacts (39Vijay SinghNo ratings yet
- 2022 Hilti Product Catalogue Firestop PDFDocument32 pages2022 Hilti Product Catalogue Firestop PDFecjr consNo ratings yet
- Requirements for Polymer MaterialsDocument4 pagesRequirements for Polymer MaterialsadrianNo ratings yet
- Development of An Internationally Competitive Australian Saffron IndustryDocument37 pagesDevelopment of An Internationally Competitive Australian Saffron IndustryNemanja NikolicNo ratings yet
- Spe 200369 PaDocument13 pagesSpe 200369 PaYousif IraqiNo ratings yet
- Energy Analysis of The Steel Making IndustryDocument7 pagesEnergy Analysis of The Steel Making Industrymrshami7754No ratings yet