You might also like
- MammalsFrom EverandMammalsG. Causey WhittowNo ratings yet
- Evaluating Thermoregulation in ReptilesDocument10 pagesEvaluating Thermoregulation in ReptilesSantiago SuarezNo ratings yet
- Extreme Measures: The Ecological Energetics of Birds and MammalsFrom EverandExtreme Measures: The Ecological Energetics of Birds and MammalsNo ratings yet
- Insect ThermoregulationDocument5 pagesInsect ThermoregulationMichaèl Kellett100% (2)
- Fires of Life: Endothermy in Birds and MammalsFrom EverandFires of Life: Endothermy in Birds and MammalsRating: 4 out of 5 stars4/5 (1)
- Energy Saving Processes in Huddling Emperor PenguiDocument9 pagesEnergy Saving Processes in Huddling Emperor PenguiLulu Nur FajriNo ratings yet
- Tattersall2016 Seasonal Endothermy ReptilDocument8 pagesTattersall2016 Seasonal Endothermy ReptilAlex RodriguezNo ratings yet
- Department of Zoology, University of Washington, Seattle, U.S.ADocument34 pagesDepartment of Zoology, University of Washington, Seattle, U.S.AUber Schalke Rozo GarciaNo ratings yet
- Rep-Jrf 114 2 001Document16 pagesRep-Jrf 114 2 001siyopin173No ratings yet
- Gómez AlesETAL2017Document10 pagesGómez AlesETAL2017laspiursaurusNo ratings yet
- GekoDocument22 pagesGekoVíctor MundoNo ratings yet
- Adaptive thermoregulation strategies in different animal speciesDocument5 pagesAdaptive thermoregulation strategies in different animal speciesRio ImaniarNo ratings yet
- Thermoregulation in BatsDocument45 pagesThermoregulation in BatsBaraa AkeelNo ratings yet
- Griebeler & Werner 2018 - Formal Comment On Myhrvold (2016) Dinosaur Metabolism and The Allometry of Maximum Growth Rate. PLoS ONE 11 (11) E0163205Document18 pagesGriebeler & Werner 2018 - Formal Comment On Myhrvold (2016) Dinosaur Metabolism and The Allometry of Maximum Growth Rate. PLoS ONE 11 (11) E0163205DavidCernyNo ratings yet
- Lin 2005Document7 pagesLin 2005Jake Floyd G. FabianNo ratings yet
- Climate Warming LizardsDocument10 pagesClimate Warming LizardsbiologaemabNo ratings yet
- Fierro Estrada2019Document12 pagesFierro Estrada2019AimeeAdrnNo ratings yet
- Raymond B. Huey Montgomery Slatkin The Quarterly Review of Biology, Vol. 51, No. 3 (Sep., 1976), 363-384Document23 pagesRaymond B. Huey Montgomery Slatkin The Quarterly Review of Biology, Vol. 51, No. 3 (Sep., 1976), 363-384Juan Sebastian BobadillaNo ratings yet
- Ashton and Feldman 2003. BERGMANN'S RULE IN NONAVIAN REPTILES - TURTLES FOLLOW IT, LIZARDS AND SNAKE REVERSE IT PDFDocument13 pagesAshton and Feldman 2003. BERGMANN'S RULE IN NONAVIAN REPTILES - TURTLES FOLLOW IT, LIZARDS AND SNAKE REVERSE IT PDFFred LimaNo ratings yet
- Manuscript SubmitDocument23 pagesManuscript Submitapi-295124915No ratings yet
- A Body Temperature Model For LizardsDocument9 pagesA Body Temperature Model For LizardsCésar MolinaNo ratings yet
- Physiological Mechanisms of Thermoregulation in ReptilesDocument9 pagesPhysiological Mechanisms of Thermoregulation in ReptilesSalvador ReyesNo ratings yet
- Physrev 1921 1 2 295 PDFDocument32 pagesPhysrev 1921 1 2 295 PDFPuru SaxenaNo ratings yet
- Blouin Demers2005Document7 pagesBlouin Demers2005Rafael Loaiza RamírezNo ratings yet
- Diurnal Variation of Heat Intake in Ovariectomized, Steroid-Treated RatsDocument11 pagesDiurnal Variation of Heat Intake in Ovariectomized, Steroid-Treated RatsFrontiersNo ratings yet
- Brood Production Increases When Artificial Heating Is Provided To Colonies of Stingless BeesDocument6 pagesBrood Production Increases When Artificial Heating Is Provided To Colonies of Stingless BeesOtaviano Carneiro Da Cunha NetoNo ratings yet
- Body Temperatures in Free - Ying PigeonsDocument5 pagesBody Temperatures in Free - Ying PigeonsDummy DonNo ratings yet
- Heat Stress CattleDocument10 pagesHeat Stress CattleRaj CellaNo ratings yet
- Rea T Transfer in Biomedical Engineering and Biotechnology: Proceedings of TheDocument13 pagesRea T Transfer in Biomedical Engineering and Biotechnology: Proceedings of TheHussain Amir AliNo ratings yet
- Avila & Medelin. 2004. Ecological, Taxonomic, and Physiological Correlates of Cave Use by Mexican Bats PDFDocument14 pagesAvila & Medelin. 2004. Ecological, Taxonomic, and Physiological Correlates of Cave Use by Mexican Bats PDFHenry CondoriNo ratings yet
- Thermal physiology of three small African mammalsDocument8 pagesThermal physiology of three small African mammalsNita SofianaNo ratings yet
- Eaax9361 FullDocument11 pagesEaax9361 FullDobby ElfoNo ratings yet
- Body Temperature Influences LifespanDocument15 pagesBody Temperature Influences LifespanNomalingeNo ratings yet
- Dependencia Térmica de La Salamandra Endémica de Colombia: Bolitoglossa Ramosi (Caudata, Plethodontidae)Document5 pagesDependencia Térmica de La Salamandra Endémica de Colombia: Bolitoglossa Ramosi (Caudata, Plethodontidae)Jehú Velásquez MoralesNo ratings yet
- Journal - Ant Thermal Tolerance a Review of Methods, Hypotheses,Document11 pagesJournal - Ant Thermal Tolerance a Review of Methods, Hypotheses,songguan26No ratings yet
- Physiological Reviews: MAX KleiberDocument31 pagesPhysiological Reviews: MAX KleiberFetcu AndreiNo ratings yet
- tmpF8C TMPDocument9 pagestmpF8C TMPFrontiersNo ratings yet
- Artigo Brahman adaptativo produtivo 2020Document7 pagesArtigo Brahman adaptativo produtivo 2020JRDRVNo ratings yet
- Heldmaier Et Al 1989 Photoperiod and Thermoregulation in Vertebrates Body Temperature Rhythms and ThermogenicDocument15 pagesHeldmaier Et Al 1989 Photoperiod and Thermoregulation in Vertebrates Body Temperature Rhythms and Thermogenicsaryasingh253No ratings yet
- PDF FuctEcol2004Document7 pagesPDF FuctEcol2004maguitcoolNo ratings yet
- Lolli 2009Document9 pagesLolli 2009Mumin HilmanNo ratings yet
- KRISTINEKRUGERR PRINZINGERK - L SCHUCHMANNCompBiochemPhysiol73A1982Document12 pagesKRISTINEKRUGERR PRINZINGERK - L SCHUCHMANNCompBiochemPhysiol73A1982Daniela GonzalezNo ratings yet
- 2008 Miyatake Et Al PHEDocument7 pages2008 Miyatake Et Al PHEJosé Augusto RoxinolNo ratings yet
- Vollet-Neto Et Al 2015Document11 pagesVollet-Neto Et Al 2015Otaviano Carneiro Da Cunha NetoNo ratings yet
- PDF-Williams Et Al 2007 ShorebirdDocument14 pagesPDF-Williams Et Al 2007 ShorebirdmaguitcoolNo ratings yet
- Animal Housing in Hot Climates PDFDocument122 pagesAnimal Housing in Hot Climates PDFIlyas FOxeérNo ratings yet
- 1 s2.0 S1751731112001838 MainDocument6 pages1 s2.0 S1751731112001838 Mainmariafernandaalves905No ratings yet
- First Page PDFDocument1 pageFirst Page PDFSumeyyah KemalNo ratings yet
- Gaughan 2007Document10 pagesGaughan 2007VALERIA DEL PILAR PEÑARANDA RINCONNo ratings yet
- Thermal Time - Concepts and Utility: Annals of Applied Biology (2005), 146:1-14Document14 pagesThermal Time - Concepts and Utility: Annals of Applied Biology (2005), 146:1-14jocyeoNo ratings yet
- Physiological and Behavioral Effects of Heat Stress in Dairy CowsDocument20 pagesPhysiological and Behavioral Effects of Heat Stress in Dairy CowsIngridAlvarezNo ratings yet
- TMP F2 DCDocument10 pagesTMP F2 DCFrontiersNo ratings yet
- Physiological Bases of Mosquito EcologyDocument11 pagesPhysiological Bases of Mosquito Ecologyreyzi hananNo ratings yet
- TsiDocument320 pagesTsiLeonides Bernardo Fernández LicónNo ratings yet
- Revista Brasileira de Zootecnia: Tami M. Brown-BrandlDocument9 pagesRevista Brasileira de Zootecnia: Tami M. Brown-BrandlyutefupNo ratings yet
- 1 Magrath 05Document15 pages1 Magrath 05Mohammed KouidriNo ratings yet
- 0022-2585 (2006) 43 (1276:eotodo) 2 0 Co 2Document12 pages0022-2585 (2006) 43 (1276:eotodo) 2 0 Co 2Luisa MatzkNo ratings yet
- On The Thermal Husbandry of Monitor Lizards: Herpetological Review December 2014Document15 pagesOn The Thermal Husbandry of Monitor Lizards: Herpetological Review December 2014Ljón BjörnNo ratings yet
- Annurev Ecolsys 37 091305Document35 pagesAnnurev Ecolsys 37 091305biofurgNo ratings yet
- Accepted Manuscript: Research in Veterinary ScienceDocument37 pagesAccepted Manuscript: Research in Veterinary ScienceWilliam EdwardsNo ratings yet
- CDI Blood Parameter Monitoring System 550: Real-Time Monitoring of DO As One of 12 Key Blood ParametersDocument5 pagesCDI Blood Parameter Monitoring System 550: Real-Time Monitoring of DO As One of 12 Key Blood ParametersManigandan DhamodhiranNo ratings yet
- Physiological Principles of SotaiDocument13 pagesPhysiological Principles of SotaiBuyu IkkanNo ratings yet
- Manonmaniam Sundaranar University: B.Sc. Psychology - I YearDocument72 pagesManonmaniam Sundaranar University: B.Sc. Psychology - I Yearhimansu reddyNo ratings yet
- Propozitii Scoase Din Cambridge Advanced DictionaryDocument369 pagesPropozitii Scoase Din Cambridge Advanced DictionaryFlorentinoTNo ratings yet
- Exercise 3 RBC CountDocument6 pagesExercise 3 RBC CountJohn Paul DianNo ratings yet
- Gene - and Cell-Based Treatment Strategies For The EyeDocument101 pagesGene - and Cell-Based Treatment Strategies For The EyeCantika MutiaraNo ratings yet
- 3 Principles of Cell MetabolismDocument53 pages3 Principles of Cell MetabolismKristine Claire TarucNo ratings yet
- English - On Farm Post Mortem Guide For BroilersDocument72 pagesEnglish - On Farm Post Mortem Guide For BroilersMahjoubNo ratings yet
- L11 - Practice Q - HBDocument4 pagesL11 - Practice Q - HBEvan YEUNG [09N12]No ratings yet
- ProteaseDocument18 pagesProteasePriyaNo ratings yet
- Aakash Model Test Papers Solutions XI Half Biology PDFDocument26 pagesAakash Model Test Papers Solutions XI Half Biology PDFSuguna RajaguruNo ratings yet
- Lecture Learning Objectives for BIO 210 SystemsDocument14 pagesLecture Learning Objectives for BIO 210 SystemsAmber DavisNo ratings yet
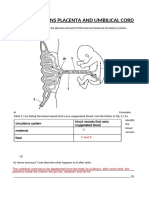
- Igcse Questions Placenta and Umbilical CordDocument3 pagesIgcse Questions Placenta and Umbilical CordFabriV9No ratings yet
- Pregnancy Induced Hypertension Nursing Care PlanDocument5 pagesPregnancy Induced Hypertension Nursing Care Planjohncarlo ramosNo ratings yet
- FrogDocument28 pagesFrogSujay RankNo ratings yet
- Phylum Echinodermata PresentationDocument38 pagesPhylum Echinodermata PresentationAliah CanalsNo ratings yet
- Annexure-97. (B.sc. (H) Botany)Document186 pagesAnnexure-97. (B.sc. (H) Botany)Sourabh RajputNo ratings yet
- (The Natural History of The Crustacea) Les Watling, Martin Thiel (Eds.) - Functional Morphology and Diversity. 1-Oxford University Press (2013)Document515 pages(The Natural History of The Crustacea) Les Watling, Martin Thiel (Eds.) - Functional Morphology and Diversity. 1-Oxford University Press (2013)Yamaly Barragán MarínNo ratings yet
- Anesthetics Drugs - ADocument72 pagesAnesthetics Drugs - Agech yayehNo ratings yet
- What Is General Adaptation SyndromeDocument3 pagesWhat Is General Adaptation SyndromeCARLOS, Ryan CholoNo ratings yet
- Advanced Human Physiology Lab ManualDocument58 pagesAdvanced Human Physiology Lab ManualRishavNo ratings yet
- Photosynthesis Basics, History and ModellingDocument27 pagesPhotosynthesis Basics, History and ModellingDario DiosNo ratings yet
- 3.1 - The Digestive SystemDocument21 pages3.1 - The Digestive SystemJavierNo ratings yet
- LAS 4 - Plant and Animal CellsDocument7 pagesLAS 4 - Plant and Animal CellsJeanne RanielleNo ratings yet
- Boardworks A2 Biology ContentsDocument16 pagesBoardworks A2 Biology ContentsStudent 365No ratings yet
- Donepezil: DR Rashmi SurtiDocument55 pagesDonepezil: DR Rashmi SurtirsurtiNo ratings yet
- Final Thesis HennaDocument127 pagesFinal Thesis HennaHenna TkNo ratings yet
- Digestive System LabsheetDocument4 pagesDigestive System LabsheetKATHLEEN MAE HERMONo ratings yet
- 14-16 Juli 2022, Jayapura I Post Test (Jawaban)Document10 pages14-16 Juli 2022, Jayapura I Post Test (Jawaban)Abdul Charis KonorasNo ratings yet
- Life Processes Notes (Yashvi Modi)Document15 pagesLife Processes Notes (Yashvi Modi)YASHVI MODI60% (5)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Why We Sleep: Unlocking the Power of Sleep and DreamsFrom EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsRating: 4.5 out of 5 stars4.5/5 (2083)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- Lymph & Longevity: The Untapped Secret to HealthFrom EverandLymph & Longevity: The Untapped Secret to HealthRating: 4.5 out of 5 stars4.5/5 (13)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)