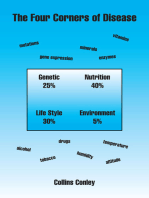
You might also like
- Fast Facts: Inflammatory Bowel Disease: Translating the science into compassionate IBD careFrom EverandFast Facts: Inflammatory Bowel Disease: Translating the science into compassionate IBD careNo ratings yet
- Clinical Risks Associated WithDocument6 pagesClinical Risks Associated WithLauura Cristina BarriosNo ratings yet
- 649 FullDocument6 pages649 FullDumitru BiniucNo ratings yet
- New Mechanisms and The Anti-Inflammatory Role of Curcumin in Obesity and Obesity-Related Metabolic DiseasesDocument11 pagesNew Mechanisms and The Anti-Inflammatory Role of Curcumin in Obesity and Obesity-Related Metabolic DiseasessudirmanahdiNo ratings yet
- Equine Metabolic SyndromeDocument7 pagesEquine Metabolic SyndromeSebastianSierraNo ratings yet
- Healthy Aging JournalDocument5 pagesHealthy Aging JournalDwi Annur PuspitaNo ratings yet
- 20.anorexia Nervosa COVID-19 Pandemic Period (Review)Document5 pages20.anorexia Nervosa COVID-19 Pandemic Period (Review)jacopo pruccoliNo ratings yet
- News Mediterranean Diet vs. Metabolic Syndrome: 1-0Document2 pagesNews Mediterranean Diet vs. Metabolic Syndrome: 1-0JasminaVejnovicNo ratings yet
- Obesity Pathophysiology and Management: Genes, Environment, and Treatment OptionsDocument26 pagesObesity Pathophysiology and Management: Genes, Environment, and Treatment OptionstiffanyNo ratings yet
- Public Health Chapter 5 ObesityDocument20 pagesPublic Health Chapter 5 ObesityQuốc Huy PhanNo ratings yet
- Canaries in The Coal Mine ArticleDocument7 pagesCanaries in The Coal Mine ArticlevibhutiNo ratings yet
- Vegetables Containing Phytochemicals With Potential Anti-ObesityDocument47 pagesVegetables Containing Phytochemicals With Potential Anti-ObesityMano RatyiNo ratings yet
- Ijbsv07p1003 PDFDocument13 pagesIjbsv07p1003 PDFDeedee RenovaldiNo ratings yet
- Obesity: A Puzzling CVD Risk FactorDocument48 pagesObesity: A Puzzling CVD Risk FactorRian Sanjaya100% (1)
- Mehu131 U1 T6 Obesidad4Document16 pagesMehu131 U1 T6 Obesidad4Valeria Moretto VegaNo ratings yet
- The China Study PDFDocument7 pagesThe China Study PDFPatrice108365No ratings yet
- Aging Adiposity and Calorie Restriction Fontana2007Document9 pagesAging Adiposity and Calorie Restriction Fontana2007rhabarberrhabarberNo ratings yet
- The Oxford Vegetarian Study: An Overview: Paul N Appleby, Margaret Thorogood, Jim I Mann, and Timothy JA KeyDocument7 pagesThe Oxford Vegetarian Study: An Overview: Paul N Appleby, Margaret Thorogood, Jim I Mann, and Timothy JA KeynikhilNo ratings yet
- Denyer 2021Document9 pagesDenyer 2021marparolaNo ratings yet
- Eaton, Konner, Cordain 2010 AJCNDocument3 pagesEaton, Konner, Cordain 2010 AJCNBarnettLudwigNo ratings yet
- An epidemiological study of environmental factors associated with canine obesityDocument6 pagesAn epidemiological study of environmental factors associated with canine obesityvetdomeupetNo ratings yet
- Available Online Through: Malnutrition Leading To Cancer by Some Environmental HazardsDocument5 pagesAvailable Online Through: Malnutrition Leading To Cancer by Some Environmental HazardsmilindNo ratings yet
- Effect of Restriction Vegan Diet's On Muscle Mass, Oxidative Status, and Myocytes Differentiation - Pilot StudyDocument9 pagesEffect of Restriction Vegan Diet's On Muscle Mass, Oxidative Status, and Myocytes Differentiation - Pilot StudyKrissy SuNo ratings yet
- Review: Saint James School of Medicine, Albert Lake Drive, The Quarter A-1 2640, Anguilla, British West IndiesDocument15 pagesReview: Saint James School of Medicine, Albert Lake Drive, The Quarter A-1 2640, Anguilla, British West IndieslintangsrNo ratings yet
- 10 1016@j Jacc 2017 11 011Document16 pages10 1016@j Jacc 2017 11 011Anita YolandhaNo ratings yet
- From Table To Stable A Comparative Review of Selected Aspects Ofhuman and Equine Metabolic SyndromeDocument8 pagesFrom Table To Stable A Comparative Review of Selected Aspects Ofhuman and Equine Metabolic SyndromeSebastianSierraNo ratings yet
- Obesity The Integrated Roles of Environment and GeneticsDocument16 pagesObesity The Integrated Roles of Environment and GeneticsEdgardo100% (1)
- Article in Press: Endocrine Disruptors: New Players in The Pathophysiology of Type 2 Diabetes?Document9 pagesArticle in Press: Endocrine Disruptors: New Players in The Pathophysiology of Type 2 Diabetes?spadini putriNo ratings yet
- Editorial: James L. Oschman, PHDDocument4 pagesEditorial: James L. Oschman, PHDKaycsa AdrianaNo ratings yet
- Editorial: James L. Oschman, PHDDocument5 pagesEditorial: James L. Oschman, PHDSofia marisa fernandesNo ratings yet
- Nutrigenomics and Nutrigenetics in Obesity and CVDsDocument50 pagesNutrigenomics and Nutrigenetics in Obesity and CVDsYair BuenrostroNo ratings yet
- Research: Persistent Organic Pollutant Exposure Leads To Insulin Resistance SyndromeDocument7 pagesResearch: Persistent Organic Pollutant Exposure Leads To Insulin Resistance SyndromeelgorilagrandeNo ratings yet
- 1 s2.0 S0092867415001865 Main PDFDocument13 pages1 s2.0 S0092867415001865 Main PDFtlidiaNo ratings yet
- Mental Consequences of ObesityDocument7 pagesMental Consequences of Obesitykeshav singhNo ratings yet
- Adipose and Skeletal Muscle Thermogenesis: Studies From Large AnimalsDocument17 pagesAdipose and Skeletal Muscle Thermogenesis: Studies From Large AnimalsBernard SoumaNo ratings yet
- Adiponectin ReviewDocument5 pagesAdiponectin ReviewAisha MurtazaNo ratings yet
- Bluher (2009) Adipose Tissue Dysfunction in ObesityDocument10 pagesBluher (2009) Adipose Tissue Dysfunction in ObesityBruno TeixeiraNo ratings yet
- Nutrigenomics Research: A ReviewDocument14 pagesNutrigenomics Research: A ReviewpuspitaNo ratings yet
- Research Quarterly explores evolutionary perspective on inactivity, obesity and diabetesDocument9 pagesResearch Quarterly explores evolutionary perspective on inactivity, obesity and diabetesRuel CedenoNo ratings yet
- Am J Clin Nutr 2015 Virtanen 1088 96Document9 pagesAm J Clin Nutr 2015 Virtanen 1088 96Marko BrzakNo ratings yet
- Egg Consumption and Risk of Incident Type 2 Diabetes in Men: The Kuopio Ischaemic Heart Disease Risk Factor StudyDocument9 pagesEgg Consumption and Risk of Incident Type 2 Diabetes in Men: The Kuopio Ischaemic Heart Disease Risk Factor StudyMatías RamírezNo ratings yet
- Obesity, Inflammation, and Periodontal Disease: Critical Reviews in Oral Biology & MedicineDocument11 pagesObesity, Inflammation, and Periodontal Disease: Critical Reviews in Oral Biology & MedicineClaudia Patricia Ballen GarzonNo ratings yet
- Zhang 2017Document25 pagesZhang 2017hayNo ratings yet
- Effects of Surgical Sterilization On Canine and Feline Health and On SocietyDocument9 pagesEffects of Surgical Sterilization On Canine and Feline Health and On SocietyNadhim Jawad HasanNo ratings yet
- Control of The Risk of Human Toxoplasmosis Transmitted by MeatDocument12 pagesControl of The Risk of Human Toxoplasmosis Transmitted by Meatreddemar7082No ratings yet
- Fuente de MetodosDocument9 pagesFuente de MetodosNinfa Margarita Alejo MoralesNo ratings yet
- Healthy Aging Diets: Focus on the Okinawan DietDocument38 pagesHealthy Aging Diets: Focus on the Okinawan DietLiana LianaNo ratings yet
- Lifestyle and Nutritional ImbalancesDocument19 pagesLifestyle and Nutritional Imbalanceskiko arrojasNo ratings yet
- HPA Axis Role in Central Obesity and Metabolic DiseasesDocument9 pagesHPA Axis Role in Central Obesity and Metabolic DiseasesDavidson De AraújoNo ratings yet
- Fuente de Metodos 3Document8 pagesFuente de Metodos 3Ninfa Margarita Alejo MoralesNo ratings yet
- Vegetarian Diets A Way Towards A Sustainable SocietyDocument15 pagesVegetarian Diets A Way Towards A Sustainable SocietyLê Thiện TrungNo ratings yet
- The Effects of A Vegan Diet On Human Health, The Environment, and Animal Welfare As Compared To A Traditional Omnivorous DietDocument13 pagesThe Effects of A Vegan Diet On Human Health, The Environment, and Animal Welfare As Compared To A Traditional Omnivorous DietLow Beng KiatNo ratings yet
- Obesity and Dementia: Adipokines Interact With The BrainDocument18 pagesObesity and Dementia: Adipokines Interact With The BrainMirian RochaNo ratings yet
- Edrv 0096Document21 pagesEdrv 0096inunk xiNo ratings yet
- Animal Fat Bact To TableDocument25 pagesAnimal Fat Bact To Tablelokesh tiwariNo ratings yet
- Rodent Models of Type 2 Diabetes MellitusDocument26 pagesRodent Models of Type 2 Diabetes MellitusAhmed khaledNo ratings yet
- Modul LLL Topic 36.1Document9 pagesModul LLL Topic 36.1Akreditasi Sungai AbangNo ratings yet
- PHD Thesis: University of Medicine and Pharmacy CraiovaDocument10 pagesPHD Thesis: University of Medicine and Pharmacy CraiovapaulNo ratings yet
- Adipose Tissue Plasticity ReviewDocument28 pagesAdipose Tissue Plasticity Reviewjonsmth552No ratings yet
- Definicion de ObesidadDocument16 pagesDefinicion de ObesidadRomina Belen Soto MoralesNo ratings yet
- How The First Nine Months Shape The Rest of Your LifeDocument9 pagesHow The First Nine Months Shape The Rest of Your Lifeمحمد عريف حكيم60% (5)
- Writing Am Not Agreeing With Everything: Thoughts. Statikus Ige: I Don't Agree Totally WithDocument7 pagesWriting Am Not Agreeing With Everything: Thoughts. Statikus Ige: I Don't Agree Totally WithannaszekerNo ratings yet
- Materi Fatty Liver Rsu Hermina & PLNDocument15 pagesMateri Fatty Liver Rsu Hermina & PLNifa virdiyasNo ratings yet
- Lordosis: Mark Anthony O.BernabeDocument11 pagesLordosis: Mark Anthony O.BernabeJayson King Legaspi CruzNo ratings yet
- AquaticExerciseDocument25 pagesAquaticExercisegialinu100% (1)
- Parenteral Nutrition CalculationsDocument62 pagesParenteral Nutrition Calculationsmikosoft2010No ratings yet
- Vildagliptin A New Oral Treatment For Type 2Document12 pagesVildagliptin A New Oral Treatment For Type 2Andhi Fahrurroji100% (1)
- Personal and Environmental WellnessDocument19 pagesPersonal and Environmental WellnessBart VillanuevaNo ratings yet
- PDP Learning Activity LogDocument3 pagesPDP Learning Activity Logapi-370434100No ratings yet
- Makalah KesehatanDocument9 pagesMakalah KesehatanNini RahmiNo ratings yet
- Breastfeeding Benefits for Mothers and BabiesDocument3 pagesBreastfeeding Benefits for Mothers and BabiesKramSenarevolNo ratings yet
- DKA FinalDocument66 pagesDKA FinalllaychinNo ratings yet
- Interpretations of Test Results in Routine Urine AnalysisDocument2 pagesInterpretations of Test Results in Routine Urine AnalysisDevaughn Ray PacialNo ratings yet
- Dieting Cheat Sheet Coach KyleDocument2 pagesDieting Cheat Sheet Coach KyleMarcusNo ratings yet
- Medical Declaration Form SummaryDocument1 pageMedical Declaration Form SummaryRamNo ratings yet
- Nursing Care Plan For Client Type 2 DM: by Fitratunnisa Harni Septria FizaDocument18 pagesNursing Care Plan For Client Type 2 DM: by Fitratunnisa Harni Septria Fiza'Ilham Malda'0% (1)
- Souvenir Program - APSAVD 2018Document36 pagesSouvenir Program - APSAVD 2018Tiel NoembreuxNo ratings yet
- 10 Things To Know About Body FatDocument1 page10 Things To Know About Body FatHilde CampNo ratings yet
- Name: Reycel Ann L. Ilawan Section: F Reference: Puhl, R., Rubino, F., Cummings, D., Eckel, R., & Ryan, D. (2020) - Joint InternationalDocument3 pagesName: Reycel Ann L. Ilawan Section: F Reference: Puhl, R., Rubino, F., Cummings, D., Eckel, R., & Ryan, D. (2020) - Joint InternationalReycel IlawanNo ratings yet
- 10 Management of Patient With Hypoglycemic Coma. Management of Patient With Hyperglycemic (Ketoacidotic) ComaDocument59 pages10 Management of Patient With Hypoglycemic Coma. Management of Patient With Hyperglycemic (Ketoacidotic) ComaMimi ChukwuNo ratings yet
- Pink Panther - Diabetes Management - Chapter 11Document18 pagesPink Panther - Diabetes Management - Chapter 11jennmoyerNo ratings yet
- MS Critical Thinking DrillsDocument18 pagesMS Critical Thinking DrillsSTEFFI GABRIELLE GOLEZ100% (1)
- Resource Sheet 3 2 5Document4 pagesResource Sheet 3 2 5api-335266803No ratings yet
- Diagnostic Test Natural Science 4º Primaria BymeDocument2 pagesDiagnostic Test Natural Science 4º Primaria BymeAntonio Javier Navarro60% (5)
- Lipo Contrast Cryo BrochureDocument5 pagesLipo Contrast Cryo BrochureSerenity ZhangNo ratings yet
- World's Best Weight Loss and Health Program Alive GuideDocument138 pagesWorld's Best Weight Loss and Health Program Alive GuideJed Diamond100% (4)
- Christian Thibaudau - HSS-100Document55 pagesChristian Thibaudau - HSS-100JesseKentNo ratings yet
- Answer Key - Epi Midterm Study Guide - 2018Document8 pagesAnswer Key - Epi Midterm Study Guide - 2018Sanaz NiksefatNo ratings yet
- Exercice TRX PDFDocument1 pageExercice TRX PDFnoreadil0% (2)