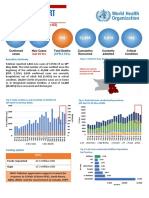
You might also like
- Vital Signs Graphic SheetDocument6 pagesVital Signs Graphic SheetMacen OrtegaNo ratings yet
- DAP Plant Optimization by JacobsDocument16 pagesDAP Plant Optimization by JacobsVinh Do ThanhNo ratings yet
- Reaction Kinetics: The Essentials: Experiment No. 2Document7 pagesReaction Kinetics: The Essentials: Experiment No. 2JV MandigmaNo ratings yet
- Bhandari1999 PDFDocument9 pagesBhandari1999 PDFMd Mahmudul HasanNo ratings yet
- Environmental Requirements: 4.2 TemperatureDocument21 pagesEnvironmental Requirements: 4.2 TemperatureJuan Camilo SantillanNo ratings yet
- E Ect of Oligomeric or Polymeric Additives On Glass Transition, Viscosity and Crystallization of Amorphous IsomaltDocument11 pagesE Ect of Oligomeric or Polymeric Additives On Glass Transition, Viscosity and Crystallization of Amorphous IsomaltSina pakNo ratings yet
- Mapping QTL For Cold Tolerance Trait in A Gift Derived Tilapia Line by Ddrad Seq Chun Hui Ai Full ChapterDocument29 pagesMapping QTL For Cold Tolerance Trait in A Gift Derived Tilapia Line by Ddrad Seq Chun Hui Ai Full Chapternancy.cloutier411100% (5)
- Articulo Agroindustrial 2Document9 pagesArticulo Agroindustrial 2Pablo GomezNo ratings yet
- 08.influence of Time Temperature Moisture Ingredients and Processing Conditions On Starch GelatinizationDocument26 pages08.influence of Time Temperature Moisture Ingredients and Processing Conditions On Starch GelatinizationKiefer YuNo ratings yet
- International Journal of Pharmaceutics: Rodrigo Soto, Michael SV ArdDocument12 pagesInternational Journal of Pharmaceutics: Rodrigo Soto, Michael SV ArdStefania ArdeleanuNo ratings yet
- Maillard Reaction of Lactose and Fluoxentine HCL A Sec AmineDocument9 pagesMaillard Reaction of Lactose and Fluoxentine HCL A Sec AmineelvaNo ratings yet
- Food Chemistry: Yeting Wu, Youyou Lu, Yuhao Huang, Hong Lin, Guanzhi Chen, Yan Chen, Zhenxing LiDocument10 pagesFood Chemistry: Yeting Wu, Youyou Lu, Yuhao Huang, Hong Lin, Guanzhi Chen, Yan Chen, Zhenxing LiZahrah KhaeraniNo ratings yet
- Taylor Et Al 2007Document12 pagesTaylor Et Al 2007animoysonrieNo ratings yet
- Effect of Different Drying Temperatures On The Moisture and Phytochemical Constituents of Edible Irish Brown SeaweedDocument7 pagesEffect of Different Drying Temperatures On The Moisture and Phytochemical Constituents of Edible Irish Brown SeaweedRosmery Naupari AlvarezNo ratings yet
- A Critical Review of Some Non-Equilibrium Situations and Glass Transitions On Water Activity Values of Foods in The Microbiological Growth RangeDocument22 pagesA Critical Review of Some Non-Equilibrium Situations and Glass Transitions On Water Activity Values of Foods in The Microbiological Growth RangeLuis GallardoNo ratings yet
- 105 Water Sorption Isotherms and Phase Transitions of Sodium Caseinate-Lipid Films AsDocument8 pages105 Water Sorption Isotherms and Phase Transitions of Sodium Caseinate-Lipid Films AsNicolas AvendañoNo ratings yet
- Review: Psychrophilic Microorganisms: Challenges For LifeDocument5 pagesReview: Psychrophilic Microorganisms: Challenges For LifeayakashimajorinNo ratings yet
- Effects of Carbohydrate Crystallization On Stability of Dehydrated Foods and Ingredient FormulationsDocument9 pagesEffects of Carbohydrate Crystallization On Stability of Dehydrated Foods and Ingredient FormulationsIsra CFNo ratings yet
- 0fcfd5093e28d328a8000000 PDFDocument7 pages0fcfd5093e28d328a8000000 PDFricky hutagalungNo ratings yet
- J of Applied Polymer Sci - 2014 - Wyatt - The Effects of Solvent Polarity and Pka On The Absorption of Solvents Into PolyDocument7 pagesJ of Applied Polymer Sci - 2014 - Wyatt - The Effects of Solvent Polarity and Pka On The Absorption of Solvents Into Polyrozan islamNo ratings yet
- Galbreathetal2006EnvBiolFishthermalmax2N 3NDocument12 pagesGalbreathetal2006EnvBiolFishthermalmax2N 3Nprueba123prueba123No ratings yet
- Low-Temperature Tolerance of Nile Tilapia, Oreochromis Niloticus: Effects of Environmental and Dietary FactorsDocument12 pagesLow-Temperature Tolerance of Nile Tilapia, Oreochromis Niloticus: Effects of Environmental and Dietary FactorsGabriel Ebodio Armenta LópezNo ratings yet
- Fouling of Heat Exchangers by Dairy Fluids - A Review: B. BansalDocument9 pagesFouling of Heat Exchangers by Dairy Fluids - A Review: B. BansalfikerykNo ratings yet
- 8261 10.1016@j.jfoodeng.2019.03.011Document34 pages8261 10.1016@j.jfoodeng.2019.03.011Renaldi ArmanNo ratings yet
- Water Activity 1Document50 pagesWater Activity 1aoauaz2000No ratings yet
- Effect of High Pressure Treatment On Thermal and Rheological Properties of LentilDocument7 pagesEffect of High Pressure Treatment On Thermal and Rheological Properties of LentilElkyn BohórquezNo ratings yet
- On The Applicability of Flory-Huggins Theory To Ternary Starch-Water-Solute SystemsDocument10 pagesOn The Applicability of Flory-Huggins Theory To Ternary Starch-Water-Solute SystemsjuarsrdNo ratings yet
- 1 s2.0 S0925521414001124 MainDocument10 pages1 s2.0 S0925521414001124 MainValentinaNo ratings yet
- Comparison in Glass Transition and Enthalpy Relaxation Between Native and Gelatinized Rice StarchesDocument12 pagesComparison in Glass Transition and Enthalpy Relaxation Between Native and Gelatinized Rice StarchesJimena LealNo ratings yet
- 1991 - Roos, Karel - Plasticizing Effect of Water On Thermal Behavior and Crystallization of Amorphous Food ModelsDocument6 pages1991 - Roos, Karel - Plasticizing Effect of Water On Thermal Behavior and Crystallization of Amorphous Food ModelsRo XaNo ratings yet
- J Foodhyd 2014 02 001Document7 pagesJ Foodhyd 2014 02 001Virnanda SyafiraNo ratings yet
- Vital Et Al 2022Document16 pagesVital Et Al 2022Bruno FreireNo ratings yet
- Artículo ImportanteDocument9 pagesArtículo ImportanteunicacomotuNo ratings yet
- Bhat Et Al., (2016)Document11 pagesBhat Et Al., (2016)Guillermo Marcelo Badillo MuñozNo ratings yet
- Ballesteros and Walters. Cryobiology, 55Document9 pagesBallesteros and Walters. Cryobiology, 55Daniel BallesterosNo ratings yet
- Heat Treatment and Fruit RipeningDocument17 pagesHeat Treatment and Fruit RipeningirfanNo ratings yet
- Effect of Water Uptake On Amorphous Inulin PropertiesDocument6 pagesEffect of Water Uptake On Amorphous Inulin PropertiesAubrey AyusteNo ratings yet
- Variation in The Heat Shock Response and Its Implication For Predicting The Effect of Global Climate Change On Species' Biogeographical Distribution Ranges and Metabolic CostsDocument9 pagesVariation in The Heat Shock Response and Its Implication For Predicting The Effect of Global Climate Change On Species' Biogeographical Distribution Ranges and Metabolic CostspachriqueNo ratings yet
- A Litterature Review of PLADocument22 pagesA Litterature Review of PLAJames WangNo ratings yet
- 1 s2.0 S027869150900060X MainDocument10 pages1 s2.0 S027869150900060X MainHatnean CristianNo ratings yet
- 1 s2.0 S0144861711000142 MainDocument4 pages1 s2.0 S0144861711000142 MainMaira CoutoNo ratings yet
- Foods: Rapid Estimation of Parameters For Gelatinization of Waxy Corn StarchDocument10 pagesFoods: Rapid Estimation of Parameters For Gelatinization of Waxy Corn StarchMuhammad IlyasNo ratings yet
- Congeneric Variability in Lifespan Extension and Onset of Senescence Suggest Active Regulation of Aging in Response To Low TemperatureDocument8 pagesCongeneric Variability in Lifespan Extension and Onset of Senescence Suggest Active Regulation of Aging in Response To Low Temperatureender000No ratings yet
- Growth Daphnia Magna Under Effects Temp Food CDDocument9 pagesGrowth Daphnia Magna Under Effects Temp Food CDroraysideblancoNo ratings yet
- Articulo de AlmidonesDocument9 pagesArticulo de AlmidonesValen Ramirez HNo ratings yet
- Kumar Nussinov Biochem 2001 Mesophile Thermophile StabilityDocument14 pagesKumar Nussinov Biochem 2001 Mesophile Thermophile StabilityJames NanorNo ratings yet
- Cach and ReleaseDocument12 pagesCach and ReleaseAndreea ȘerbanNo ratings yet
- Magnetic Resonance Studies of Food FreezingDocument10 pagesMagnetic Resonance Studies of Food FreezingzheeveiNo ratings yet
- Marine Phytoplankton Functional Types Exhibit Diverse Responses To Thermal ChangeDocument9 pagesMarine Phytoplankton Functional Types Exhibit Diverse Responses To Thermal ChangeSurya PratamaNo ratings yet
- Repolho Et Al 2014 Fisiologia Embriones O Vulgaris TemperaturaDocument10 pagesRepolho Et Al 2014 Fisiologia Embriones O Vulgaris TemperaturaSadot Abimael RodriguezNo ratings yet
- Journal of Thermal Biology: A B B A ADocument6 pagesJournal of Thermal Biology: A B B A AJosue EscarcegaNo ratings yet
- Comparative Biochemistry and Physiology, Part A: A. Couto, P. Enes, H. Peres, A. Oliva-TelesDocument6 pagesComparative Biochemistry and Physiology, Part A: A. Couto, P. Enes, H. Peres, A. Oliva-TelesAntong AmiruddinNo ratings yet
- Phase Transitions and Structure of Solid Food Matrices: Yrjo RoosDocument6 pagesPhase Transitions and Structure of Solid Food Matrices: Yrjo RoosRaúl HernándezNo ratings yet
- Entropic Effects Enable Life at Extreme Temperatures: ResearcharticleDocument9 pagesEntropic Effects Enable Life at Extreme Temperatures: Researcharticlemichaeljacksonfan1No ratings yet
- Renewable and Sustainable Energy Reviews: Erqi Nie, Pinjing He, Hua Zhang, Liping Hao, Liming Shao, Fan LüDocument11 pagesRenewable and Sustainable Energy Reviews: Erqi Nie, Pinjing He, Hua Zhang, Liping Hao, Liming Shao, Fan Lücaio junqueira ferreiraNo ratings yet
- Proteomic Characterization of The Acid Tolerance Response in Lactococcus Lactis MG1363Document14 pagesProteomic Characterization of The Acid Tolerance Response in Lactococcus Lactis MG1363ThanasisNo ratings yet
- Improving The Stability of Glutamate Fermentation by Corynebacterium Glutamicum Via Supplementing Sorbitol or GlycerolDocument11 pagesImproving The Stability of Glutamate Fermentation by Corynebacterium Glutamicum Via Supplementing Sorbitol or GlycerolEzra KarosekaliNo ratings yet
- Enrichment of Ice Cream With Dietary Fibre: Effects On Rheological Properties, Ice Crystallisation and Glass Transition PhenomenaDocument8 pagesEnrichment of Ice Cream With Dietary Fibre: Effects On Rheological Properties, Ice Crystallisation and Glass Transition PhenomenaHugo SerafiniNo ratings yet
- Sensitivity of Metabolic Rate, Growth, and Fecundity of Tadpole Shrimp To Environmental VariationDocument7 pagesSensitivity of Metabolic Rate, Growth, and Fecundity of Tadpole Shrimp To Environmental Variationragweed420No ratings yet
- References CoccolithophoresDocument68 pagesReferences CoccolithophoresArt KNo ratings yet
- Salmonella Cold Stress Response - Mechanisms and Occurrence in FoodsDocument38 pagesSalmonella Cold Stress Response - Mechanisms and Occurrence in FoodsKatia RamónNo ratings yet
- Glass Transition and Phase Transitions in Food and Biological MaterialsFrom EverandGlass Transition and Phase Transitions in Food and Biological MaterialsNo ratings yet
- Metabolic Adaptation to Climate and Distribution of the Raccoon Procyon Lotor and Other ProcyonidaeFrom EverandMetabolic Adaptation to Climate and Distribution of the Raccoon Procyon Lotor and Other ProcyonidaeNo ratings yet
- Chitin Chitosan ProductionDocument19 pagesChitin Chitosan ProductionOmer MukhtarNo ratings yet
- CC 4593 enDocument20 pagesCC 4593 enOmer MukhtarNo ratings yet
- SitRep 19th May 2020 (ML)Document2 pagesSitRep 19th May 2020 (ML)Omer MukhtarNo ratings yet
- SitRep 19th May 2020 (ML)Document2 pagesSitRep 19th May 2020 (ML)Omer MukhtarNo ratings yet
- Viskositäts Standards 2018Document2 pagesViskositäts Standards 2018Omer MukhtarNo ratings yet
- Viskositäts Standards 2018Document2 pagesViskositäts Standards 2018Omer MukhtarNo ratings yet
- United States Patent (19) : 75) Inventors: Bruce W. Brown Virgil E. Copple 57Document7 pagesUnited States Patent (19) : 75) Inventors: Bruce W. Brown Virgil E. Copple 57Omer MukhtarNo ratings yet
- Inactive Dry Yeast For BreadDocument2 pagesInactive Dry Yeast For BreadOmer MukhtarNo ratings yet
- Process Analyzer Sample Systems: Home BlogDocument8 pagesProcess Analyzer Sample Systems: Home BlograhulNo ratings yet
- Polycarbonate Sheets NotesDocument7 pagesPolycarbonate Sheets NotesAdrian PerezNo ratings yet
- IGCSE Chemistry Revision NotesDocument32 pagesIGCSE Chemistry Revision NotesMary Pissaridou25% (4)
- Chemical Engineering Dissertation ExamplesDocument8 pagesChemical Engineering Dissertation ExamplesNeedHelpWithPaperErie100% (1)
- PRACTICE PROBLEMS - Precipitation TitrationDocument2 pagesPRACTICE PROBLEMS - Precipitation TitrationKwien Austria100% (1)
- GP 06-40 - Pipeline Coating SelectionDocument17 pagesGP 06-40 - Pipeline Coating SelectionTeymur RegenmaisterNo ratings yet
- Water Quality For FishDocument2 pagesWater Quality For FishzulfiqaralimalikNo ratings yet
- HAC-10143-MS-CIV-003-Method Statement For Concrete RepairDocument26 pagesHAC-10143-MS-CIV-003-Method Statement For Concrete RepairHema playsNo ratings yet
- IntroductionDocument15 pagesIntroductionDeepak SahNo ratings yet
- Oilfield Products - Valves & Wellheads: YyycwuvtcnkcprkrgnkpgxcnxgeqocwDocument48 pagesOilfield Products - Valves & Wellheads: Yyycwuvtcnkcprkrgnkpgxcnxgeqocwjhonny barrantesNo ratings yet
- Utilities Used in Textile Industry: University of Karachi Chemical EngineeringDocument27 pagesUtilities Used in Textile Industry: University of Karachi Chemical EngineeringAneesha PandaNo ratings yet
- Boiling Point DataDocument283 pagesBoiling Point DataKaryshma GillNo ratings yet
- Is 1367 13 1983Document7 pagesIs 1367 13 1983Dharmendra TomarNo ratings yet
- Lower Secondary Science 7 Scheme of WorkDocument21 pagesLower Secondary Science 7 Scheme of WorksumnamundethNo ratings yet
- Polymer Additives: Product Solutions From CorbionDocument8 pagesPolymer Additives: Product Solutions From CorbionXuân Giang NguyễnNo ratings yet
- ADDMIX 215 - v2Document4 pagesADDMIX 215 - v2Ankita Baban GavadeNo ratings yet
- Missile TechnologyDocument52 pagesMissile TechnologySanjay KumarNo ratings yet
- Data Sheet Permutex WF-78-123Document1 pageData Sheet Permutex WF-78-123Mariana MeirelesNo ratings yet
- Hpi Brochure PDFDocument20 pagesHpi Brochure PDFShayma'a H. HarranNo ratings yet
- To Study The Digestion of Starch by SaliDocument12 pagesTo Study The Digestion of Starch by SaliUthaya SurianNo ratings yet
- Surfaces and Interfaces: Ratnesh Kumar Sharma, Randip Kumar Das, Shiv Ranjan KumarDocument10 pagesSurfaces and Interfaces: Ratnesh Kumar Sharma, Randip Kumar Das, Shiv Ranjan KumarYulian GalihNo ratings yet
- A263-12 Stainless Chromium Steel-Clad PlateDocument6 pagesA263-12 Stainless Chromium Steel-Clad PlatemehmetNo ratings yet
- Project Report On Oleum: Guided By: Yamini S. PatelDocument51 pagesProject Report On Oleum: Guided By: Yamini S. PatelPatel RajNo ratings yet
- Dialight Chemical CompatibilityDocument16 pagesDialight Chemical CompatibilityJuan AcuñaNo ratings yet
- N-Acetylcysteine: Reversal Agent/Antidote Toxicant/Main Indications CommentsDocument3 pagesN-Acetylcysteine: Reversal Agent/Antidote Toxicant/Main Indications CommentsAshner ShatovNo ratings yet
- A Review On PerfumeryDocument14 pagesA Review On PerfumeryKhoiNguyen1205No ratings yet
- Nitrophoska Special BrochureDocument8 pagesNitrophoska Special Brochuresree778899No ratings yet