You might also like
- Delta Opioid Receptor Pharmacology and Therapeutic ApplicationsFrom EverandDelta Opioid Receptor Pharmacology and Therapeutic ApplicationsEmily M. JutkiewiczNo ratings yet
- Pharmacology 2 NbomeDocument9 pagesPharmacology 2 NbomeDylan SmithNo ratings yet
- 5-HT4 receptor agonists enhance cognition and modulate amyloid precursor protein processingDocument11 pages5-HT4 receptor agonists enhance cognition and modulate amyloid precursor protein processingihsan fadhilahNo ratings yet
- D.M. Weiner Et Al - 5-Hydroxytryptamine2A Receptor Inverse Agonists As AntipsychoticsDocument9 pagesD.M. Weiner Et Al - 5-Hydroxytryptamine2A Receptor Inverse Agonists As AntipsychoticsCortate15gNo ratings yet
- Journal AphasiaDocument9 pagesJournal AphasiaChintya Anugrah SuhendraNo ratings yet
- 1 s2.0 S0006322397002953nlnl Nnnmain PDFDocument3 pages1 s2.0 S0006322397002953nlnl Nnnmain PDFElisa SalakayNo ratings yet
- Serotonin Function in Panic Disorder: Important, But Why?: PerspectiveDocument11 pagesSerotonin Function in Panic Disorder: Important, But Why?: PerspectiveraadNo ratings yet
- Mechanisms of Pharmacotherapies for OCDDocument12 pagesMechanisms of Pharmacotherapies for OCDjosephbaroneNo ratings yet
- N Benztl Hansen2014Document7 pagesN Benztl Hansen2014AdamNo ratings yet
- Control of Serotonergic Function in Medial Prefrontal Cortex by Serotonin-2A Receptors Through A Glutamate-Dependent MechanismDocument11 pagesControl of Serotonergic Function in Medial Prefrontal Cortex by Serotonin-2A Receptors Through A Glutamate-Dependent MechanismLonkesNo ratings yet
- 1998 - Winter, Et Al - The Paradox of 5-Methoxy-N, N-DimethyltryptamineDocument8 pages1998 - Winter, Et Al - The Paradox of 5-Methoxy-N, N-DimethyltryptamineCarloNo ratings yet
- HHS Public Access: Serotonin 2A Receptors Are A Stress Response System: Implications For Post-Traumatic Stress DisorderDocument22 pagesHHS Public Access: Serotonin 2A Receptors Are A Stress Response System: Implications For Post-Traumatic Stress DisorderDebbyNo ratings yet
- The Serotonin 5-HT Receptor Agonist TCB-2: A Behavioral and Neurophysiological AnalysisDocument12 pagesThe Serotonin 5-HT Receptor Agonist TCB-2: A Behavioral and Neurophysiological AnalysisSukh MinhasNo ratings yet
- Journal of Neurochemistry - 2016 - Jalewa - Novel Incretin Analogues Improve Autophagy and Protect From MitochondrialDocument13 pagesJournal of Neurochemistry - 2016 - Jalewa - Novel Incretin Analogues Improve Autophagy and Protect From Mitochondrialjalewa.jaishreeNo ratings yet
- European Journal of Medicinal ChemistryDocument18 pagesEuropean Journal of Medicinal Chemistrybhagwan yadavNo ratings yet
- Effects of Psilocybin On Hippocampal Neurogenesis and Extinction of Trace Fear ConditioningDocument11 pagesEffects of Psilocybin On Hippocampal Neurogenesis and Extinction of Trace Fear ConditioningurkogNo ratings yet
- Hasler2004 PDFDocument12 pagesHasler2004 PDFJosé Carlos Solís SuárezNo ratings yet
- Agonistas Inversos Sesgados 5HT2A y Psicosis 2020 DiezDocument2 pagesAgonistas Inversos Sesgados 5HT2A y Psicosis 2020 DiezLuis CarbonettiNo ratings yet
- 6578 FullDocument12 pages6578 Fullシン イNo ratings yet
- Structure-Activity Relationships of Phenylalkylamines As Agonist Ligands For 5-HT2A ReceptorsDocument11 pagesStructure-Activity Relationships of Phenylalkylamines As Agonist Ligands For 5-HT2A ReceptorsAlex FulkroadNo ratings yet
- Fphar 06 00225Document17 pagesFphar 06 00225kdkijejekNo ratings yet
- Fluoxetine Impulsive BPDDocument6 pagesFluoxetine Impulsive BPDM RCNo ratings yet
- Fu-Ming Zhou and John J. Hablitz - Activation of Serotonin Receptors Modulates Synaptic Transmission in Rat Cerebral CortexDocument12 pagesFu-Ming Zhou and John J. Hablitz - Activation of Serotonin Receptors Modulates Synaptic Transmission in Rat Cerebral CortexCortate15gNo ratings yet
- Tryptophan Research in Panic Disorder: Experimental StudiesDocument8 pagesTryptophan Research in Panic Disorder: Experimental StudiesfontnmayerNo ratings yet
- Serotonin Receptor Agonism in The Expression of Behavioral Dopaminergic Supersensitivity in Subchronic Haloperidol Treated RatsDocument10 pagesSerotonin Receptor Agonism in The Expression of Behavioral Dopaminergic Supersensitivity in Subchronic Haloperidol Treated RatsLonkesNo ratings yet
- Ref 47 hasil291096-862829963C2173Aaid-ajmg173B2-0Document5 pagesRef 47 hasil291096-862829963C2173Aaid-ajmg173B2-0Fitri anaNo ratings yet
- Effects of Serotonin Transporter Promoter Polymorphisms On Serotonin FunctionDocument9 pagesEffects of Serotonin Transporter Promoter Polymorphisms On Serotonin FunctionAnzari MuhammadNo ratings yet
- %28sici%291096-9861%2820000207%29417%3A2%3C181%3A%3Aaid-cne4%3E3.0.co%3B2-aDocument14 pages%28sici%291096-9861%2820000207%29417%3A2%3C181%3A%3Aaid-cne4%3E3.0.co%3B2-agslifirsNo ratings yet
- J.C. Winter, K.C. Rice, D.J. Amorosi and R.A. Rabin - Psilocybin-Induced Stimulus Control in The RatDocument18 pagesJ.C. Winter, K.C. Rice, D.J. Amorosi and R.A. Rabin - Psilocybin-Induced Stimulus Control in The RatJunmajNo ratings yet
- %28sici%291098-2396%2820000601%2936%3A3%3C205%3A%3Aaid-syn5%3E3.0.co%3B2-dDocument17 pages%28sici%291098-2396%2820000601%2936%3A3%3C205%3A%3Aaid-syn5%3E3.0.co%3B2-dgslifirsNo ratings yet
- Biochemical Pharmacology: SciencedirectDocument13 pagesBiochemical Pharmacology: SciencedirectAline AssunçãoNo ratings yet
- Neurotoxis Phen FenDocument7 pagesNeurotoxis Phen FendrdraufNo ratings yet
- Cannabinoid Receptor Agonists Upregulate and Enhance Serotonin 5-Ht2a Receptor Activity Via Erk1 and 2 SignalingDocument23 pagesCannabinoid Receptor Agonists Upregulate and Enhance Serotonin 5-Ht2a Receptor Activity Via Erk1 and 2 SignalingabyssnepNo ratings yet
- The in Vitro Receptor Profile of Rotigotine: A New Agent For The Treatment of ParkinsonDocument14 pagesThe in Vitro Receptor Profile of Rotigotine: A New Agent For The Treatment of ParkinsonPietro BracagliaNo ratings yet
- Nemo Da 2009Document3 pagesNemo Da 2009Merari Lugo OcañaNo ratings yet
- Oamjms 9t3 288Document6 pagesOamjms 9t3 288Dewi NofiantiNo ratings yet
- Review: in The Search For Selective Ligands of 5-Ht, 5-HT and 5-Ht Serotonin ReceptorsDocument15 pagesReview: in The Search For Selective Ligands of 5-Ht, 5-HT and 5-Ht Serotonin ReceptorsAlegría LeeNo ratings yet
- Analysis of The Serotonin Transporter Gene Linked Polymorphism (5-HTTLPR) in Anorexia NervosaDocument3 pagesAnalysis of The Serotonin Transporter Gene Linked Polymorphism (5-HTTLPR) in Anorexia NervosaGerson Moreno BermúdezNo ratings yet
- Behavioral Evidence For Interactions Between A Hallucinogenic Drug and Group II Metabotropic Glutamate ReceptorsDocument8 pagesBehavioral Evidence For Interactions Between A Hallucinogenic Drug and Group II Metabotropic Glutamate ReceptorsCortate15gNo ratings yet
- The Pathophysiology and Genetics of OCDDocument12 pagesThe Pathophysiology and Genetics of OCDCrescent FangNo ratings yet
- tmpE99B TMPDocument16 pagestmpE99B TMPFrontiersNo ratings yet
- Tugas AntidepressantDocument10 pagesTugas AntidepressantSMPU ditjenbunNo ratings yet
- 6-Hydroxybuspirone Is A Major Active Metabolite of Buspirone: Assessment of Pharmacokinetics and 5-Hydroxytryptamine Receptor Occupancy in RatsDocument6 pages6-Hydroxybuspirone Is A Major Active Metabolite of Buspirone: Assessment of Pharmacokinetics and 5-Hydroxytryptamine Receptor Occupancy in RatsT h o r y n R a m o sNo ratings yet
- Pharmacology of AutacoidsDocument13 pagesPharmacology of AutacoidsInocenteNo ratings yet
- Altered Glucocorticoid Rhythm Attenuates The Ability of A Chronic SSRI To Elevate Forebrain 5-HT: Implications For The Treatment of DepressionDocument7 pagesAltered Glucocorticoid Rhythm Attenuates The Ability of A Chronic SSRI To Elevate Forebrain 5-HT: Implications For The Treatment of DepressionCony GSNo ratings yet
- 2011 - Hutchenson, Et Al - Serotonin Receptors and Valve Heart Disease - It Was Meant 2BDocument29 pages2011 - Hutchenson, Et Al - Serotonin Receptors and Valve Heart Disease - It Was Meant 2Bcolompar80No ratings yet
- Metabolic StressDocument3 pagesMetabolic StressRoberto MartineNo ratings yet
- Ijnp Pyv045 FullDocument35 pagesIjnp Pyv045 FullNityananda PortelladaNo ratings yet
- NMDA Receptor Antagonist Effects, Cortical Glutamatergic Function, and Schizophrenia: Toward A Paradigm Shift in Medication DevelopmentDocument19 pagesNMDA Receptor Antagonist Effects, Cortical Glutamatergic Function, and Schizophrenia: Toward A Paradigm Shift in Medication DevelopmentCortate15gNo ratings yet
- Teruo Hayashi Et Al - Ca 2+ Signaling Via Sigma1-Receptors: Novel Regulatory Mechanism Affecting Intracellular Ca 2+ Concentration1Document11 pagesTeruo Hayashi Et Al - Ca 2+ Signaling Via Sigma1-Receptors: Novel Regulatory Mechanism Affecting Intracellular Ca 2+ Concentration1LonkesNo ratings yet
- Drug Metab Dispos 2005 Maher 956 62Document7 pagesDrug Metab Dispos 2005 Maher 956 62Anshari StressNo ratings yet
- Invertebrate Serotonin Receptors: A Molecular Perspective On Classification and PharmacologyDocument11 pagesInvertebrate Serotonin Receptors: A Molecular Perspective On Classification and PharmacologyRofinur NuransyahNo ratings yet
- 1 s2.0 S0028390807003450 MainDocument10 pages1 s2.0 S0028390807003450 MainBianca FurduiNo ratings yet
- Gut, Microbiome, and Brain Regulatory Axis: Relevance To Neurodegenerative and Psychiatric DisordersDocument10 pagesGut, Microbiome, and Brain Regulatory Axis: Relevance To Neurodegenerative and Psychiatric DisordersVitor PereiraNo ratings yet
- Gut, Microbiome, and Brain Regulatory Axis: Relevance To Neurodegenerative and Psychiatric DisordersDocument10 pagesGut, Microbiome, and Brain Regulatory Axis: Relevance To Neurodegenerative and Psychiatric DisordersLori EsbeltoNo ratings yet
- Pharmacogenetics in Psychiatry: Translating Research Into Clinical PracticeDocument10 pagesPharmacogenetics in Psychiatry: Translating Research Into Clinical PracticeNabila ChakourNo ratings yet
- Investigating the antidepressant effects of agmatine and allopregnanolone interaction in olfactory bulbectomized ratsDocument15 pagesInvestigating the antidepressant effects of agmatine and allopregnanolone interaction in olfactory bulbectomized ratsnikuNo ratings yet
- Common and Distinct Neural Effects of Risperidone and Olanzapine During Procedural Learning in Schizophrenia: A Randomised Longitudinal fMRI StudyDocument11 pagesCommon and Distinct Neural Effects of Risperidone and Olanzapine During Procedural Learning in Schizophrenia: A Randomised Longitudinal fMRI StudyIntan Sari UmarNo ratings yet
- Paper 1Document11 pagesPaper 1Anonymous K8fT6HNo ratings yet
- Constitutive Activation of Gas Within Forebrain Neurons Causes Deficits in Sensorimotor Gating Because of PKA-Dependent Decreases in cAMPDocument12 pagesConstitutive Activation of Gas Within Forebrain Neurons Causes Deficits in Sensorimotor Gating Because of PKA-Dependent Decreases in cAMPAGRANo ratings yet
- Prelim ncm105Document4 pagesPrelim ncm105klirt carayoNo ratings yet
- Navigating The Labyrinth of Love 2013cDocument15 pagesNavigating The Labyrinth of Love 2013cvictoriaNo ratings yet
- Nasw Code of Ethics Exam For ElearningDocument4 pagesNasw Code of Ethics Exam For Elearningapi-97308101No ratings yet
- Manual HandlingDocument27 pagesManual HandlingAtinzz ChrizNo ratings yet
- 18 Min Workout PDFDocument12 pages18 Min Workout PDFdb0003No ratings yet
- Medication Booklet and TicketDocument3 pagesMedication Booklet and TicketCayanne ChuaNo ratings yet
- Neurodegenerative Disease Among Male Elite Football (Soccer) Players in Sweden: A Cohort Study - TheDocument3 pagesNeurodegenerative Disease Among Male Elite Football (Soccer) Players in Sweden: A Cohort Study - Thepitufi11No ratings yet
- Konsep Kebidanan KomunitasDocument63 pagesKonsep Kebidanan Komunitasdewi susilawatiNo ratings yet
- Cancer Screening Call ScriptDocument2 pagesCancer Screening Call Scriptjhez tel100% (1)
- Business DemographyDocument15 pagesBusiness DemographyNeha ShuklaNo ratings yet
- Quick RemapDocument32 pagesQuick RemapmollitonyNo ratings yet
- In (Ampinex, Clovilin, Vatacil)Document2 pagesIn (Ampinex, Clovilin, Vatacil)karenmichellelecarozNo ratings yet
- 04ijpr01097 1Document7 pages04ijpr01097 1ManojNo ratings yet
- The University of Melbourne Tuition Fees 2020: Tuition Fee Tables For International StudentsDocument32 pagesThe University of Melbourne Tuition Fees 2020: Tuition Fee Tables For International StudentsRuchi ChinfhaChinthaNo ratings yet
- Chapter 6 - Adipose TissueDocument13 pagesChapter 6 - Adipose TissueREMAN ALINGASANo ratings yet
- The Hospital Noise Project: Lessons On Addressing Noise From 241 U.S. HospitalsDocument50 pagesThe Hospital Noise Project: Lessons On Addressing Noise From 241 U.S. HospitalsmanishNo ratings yet
- Principles of Paragraph Writing IIIDocument15 pagesPrinciples of Paragraph Writing IIITitiek DwiNo ratings yet
- UnitedHealthcare Claim Reconsideration Form GuideDocument1 pageUnitedHealthcare Claim Reconsideration Form GuidebcvfadsNo ratings yet
- Flow RateDocument2 pagesFlow Rateفيرمان ريشادNo ratings yet
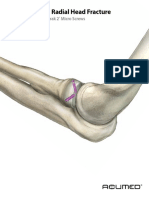
- Acutrak2 Radial Head Case StudyDocument2 pagesAcutrak2 Radial Head Case StudyDinesh KumarNo ratings yet
- Environmental Protection Act (1986)Document31 pagesEnvironmental Protection Act (1986)RUTUJA GOGULWAR100% (1)
- Ds Pedia WardDocument2 pagesDs Pedia WardRhea Mae Valles - ReyesNo ratings yet
- Safety Data Sheet Picric Acid (Alcoholic)Document14 pagesSafety Data Sheet Picric Acid (Alcoholic)tarek aneesNo ratings yet
- MELC-Based Health PresentationDocument17 pagesMELC-Based Health PresentationNelson Manalo88% (8)
- ROO-ALL-HS-PRO-0025 REV 10 Ground Disturbance and Excavation Procedure - ...Document25 pagesROO-ALL-HS-PRO-0025 REV 10 Ground Disturbance and Excavation Procedure - ...Murtadda MohammedNo ratings yet
- Capstone SolutionDocument1 pageCapstone Solutiondave magcawasNo ratings yet
- Pathophysiology of PneumoniaDocument2 pagesPathophysiology of PneumoniaJeffrey Ramos100% (1)
- Project SarvodayaDocument21 pagesProject SarvodayaArun Kumar PNo ratings yet
- H113 9 2010 9 EngDocument64 pagesH113 9 2010 9 EnguncleadolphNo ratings yet