You might also like
- Low Sugar So SimpleDocument284 pagesLow Sugar So SimpleTianur Maria100% (2)
- Diabetes Mellitus NCPDocument7 pagesDiabetes Mellitus NCPjfgnzls182892% (12)
- Case Study - DM II NewDocument77 pagesCase Study - DM II NewCraig MacaraigNo ratings yet
- Sitagliptin Slide Set Specialist - 1Document31 pagesSitagliptin Slide Set Specialist - 1Fannia Setiani100% (1)
- Experimental Biology and MedicineDocument11 pagesExperimental Biology and MedicineYunitasya GuspiraNo ratings yet
- Streptozotocin Systematic Name Chemical Data C Pharmacokinetic DataDocument2 pagesStreptozotocin Systematic Name Chemical Data C Pharmacokinetic DataNurfadilaYahyaNo ratings yet
- Literature Review: Expense ofDocument43 pagesLiterature Review: Expense ofAnn DassNo ratings yet
- Glutathione: Methods of Sample Preparation For Chromatography and Capillary ElectrophoresisDocument7 pagesGlutathione: Methods of Sample Preparation For Chromatography and Capillary ElectrophoresisBogdan CioroiuNo ratings yet
- Ganesaratnam K. Balendiran Rajesh Dabur Deborah Fraser (2004) - The Role of Glutathione in Cancer.Document10 pagesGanesaratnam K. Balendiran Rajesh Dabur Deborah Fraser (2004) - The Role of Glutathione in Cancer.Luis ReyNo ratings yet
- Tunali 2019Document5 pagesTunali 2019徳利雅No ratings yet
- Alkaloids Induce Programmed Cell Death in Bloodstream Forms of Trypanosomes (Trypanosoma B. Brucei)Document12 pagesAlkaloids Induce Programmed Cell Death in Bloodstream Forms of Trypanosomes (Trypanosoma B. Brucei)Kayo PaivaNo ratings yet
- Chemico-Biological InteractionsDocument15 pagesChemico-Biological InteractionsAamir SohailNo ratings yet
- One Systemic Protect Ant Against Oxidative and Free Radical DamageDocument19 pagesOne Systemic Protect Ant Against Oxidative and Free Radical DamageNicky ElsworthNo ratings yet
- Therapeutic Potential of Streptozotocin and It's Correlation With Human and Animal Sciences: A ReviewDocument7 pagesTherapeutic Potential of Streptozotocin and It's Correlation With Human and Animal Sciences: A ReviewIJRASETPublicationsNo ratings yet
- Rolul Taurinei in SNCDocument6 pagesRolul Taurinei in SNCMihaela ȚarnăNo ratings yet
- 6 1243 PDFDocument7 pages6 1243 PDFShawnNo ratings yet
- GSH GlutatiónDocument12 pagesGSH GlutatiónCat OsoNo ratings yet
- Agarwal Biomolecules 2019Document12 pagesAgarwal Biomolecules 2019Anil Kumar Agarwal ChemistryNo ratings yet
- Progesterone Mitigates Trimethyltin-Induced Hippocampal Damage Through Oxidative Stress BiomarkersDocument5 pagesProgesterone Mitigates Trimethyltin-Induced Hippocampal Damage Through Oxidative Stress BiomarkersomoolafolahanmiNo ratings yet
- Cleavage of Vesicular Glutamate TransportersDocument12 pagesCleavage of Vesicular Glutamate TransportersJoao GomesNo ratings yet
- J. Biol. Chem.-2002-Ma-39343-9Document8 pagesJ. Biol. Chem.-2002-Ma-39343-9Melin YohanaNo ratings yet
- Molecules 15 07235Document31 pagesMolecules 15 07235János CsernákNo ratings yet
- Bioorganic & Medicinal Chemistry LettersDocument5 pagesBioorganic & Medicinal Chemistry LettersIlham PerdanaNo ratings yet
- Stress Oksidatif DM 4Document7 pagesStress Oksidatif DM 4Almas TNo ratings yet
- ABC Transporters Glycosomal2011Document10 pagesABC Transporters Glycosomal2011franklinNo ratings yet
- Mitochondrial ApoptosisDocument5 pagesMitochondrial ApoptosisRuheen MariamNo ratings yet
- Glutation 1Document4 pagesGlutation 1Jorge Luis Plasencia CubaNo ratings yet
- O-Glcnacase Inhibitors As Potential Therapeutics For The Treatment of Alzheimer'S Disease and Related Tauopathies: Analysis of The Patent LiteratureDocument39 pagesO-Glcnacase Inhibitors As Potential Therapeutics For The Treatment of Alzheimer'S Disease and Related Tauopathies: Analysis of The Patent LiteratureHasna NoerNo ratings yet
- Base Excision Repair and CancerDocument17 pagesBase Excision Repair and CancerIonela Petronela Tarabuta BocanceaNo ratings yet
- Neurotoxicology: Zhenquan Jia, Bhaba R. Misra, Hong Zhu, Yunbo Li, Hara P. MisraDocument9 pagesNeurotoxicology: Zhenquan Jia, Bhaba R. Misra, Hong Zhu, Yunbo Li, Hara P. MisraZhenquan JiaNo ratings yet
- Menzies 2015Document13 pagesMenzies 2015kosikevinonuNo ratings yet
- Assignment On Glutathion S TransferaseDocument8 pagesAssignment On Glutathion S TransferaseSayema KhanumNo ratings yet
- Rodent Models of Streptozotocin-Induced Diabetic NephropathyDocument6 pagesRodent Models of Streptozotocin-Induced Diabetic NephropathyManishankar PampanaNo ratings yet
- Jurnal ProteaseDocument19 pagesJurnal Proteasedesmila alfnNo ratings yet
- Base Excision Repair Capacity in Mitochondria and Nuclei: Tissue-Specific VariationsDocument8 pagesBase Excision Repair Capacity in Mitochondria and Nuclei: Tissue-Specific VariationsNestor OliNo ratings yet
- Jain Et Al-2009-Protein ScienceDocument13 pagesJain Et Al-2009-Protein ScienceCelso LandichoNo ratings yet
- ArtigoDocument12 pagesArtigoGabrielNo ratings yet
- 5 PBDocument17 pages5 PBAisha MurtazaNo ratings yet
- MreC and MreD Balance The InteractionDocument39 pagesMreC and MreD Balance The InteractionVictor HugoNo ratings yet
- Controversial Impact of Sirtuins in Chronic Non-Transmissible Diseases and Rehabilitation MedicineDocument20 pagesControversial Impact of Sirtuins in Chronic Non-Transmissible Diseases and Rehabilitation MedicinerafaelorcyNo ratings yet
- 15 Glutathione Metabolism and Its Implications For HealthDocument4 pages15 Glutathione Metabolism and Its Implications For HealthTibério VasconcelosNo ratings yet
- Xue 2021Document11 pagesXue 2021Camilla RequiãoNo ratings yet
- NewInsights Into The Use of Empagliflozin-A Comprehensive ReviewDocument21 pagesNewInsights Into The Use of Empagliflozin-A Comprehensive ReviewLu MVNo ratings yet
- Glutathione and Its Role in Cellular FunctionsDocument6 pagesGlutathione and Its Role in Cellular FunctionsHoài Nam PhanNo ratings yet
- Glutathion SynthesisDocument31 pagesGlutathion SynthesisSukamto AbiyaNo ratings yet
- Estructura y Arquitectura Del PeptidoglucanoDocument19 pagesEstructura y Arquitectura Del PeptidoglucanoOleggValVilNo ratings yet
- Glutathione - WikipediaDocument75 pagesGlutathione - WikipediaKhaled MahmoudNo ratings yet
- Quercetin PankreasDocument7 pagesQuercetin PankreasAlmas TNo ratings yet
- Prostaglandin: Continued DonationsDocument27 pagesProstaglandin: Continued DonationsrajeevmkNo ratings yet
- Role of ROS in Metabolic Diseases and ChronicDocument9 pagesRole of ROS in Metabolic Diseases and ChronicFian AldyNo ratings yet
- Li Et Al. - 2009 - Novel Inhibitors Complexed With Glutamate Dehydrogenase Allosteric Regulation by Control of Protein DynamicsDocument14 pagesLi Et Al. - 2009 - Novel Inhibitors Complexed With Glutamate Dehydrogenase Allosteric Regulation by Control of Protein DynamicsErikNo ratings yet
- DiacylglycerolDocument11 pagesDiacylglycerolMai NabiehNo ratings yet
- Pro Tea SomeDocument7 pagesPro Tea SomeJuan Garzón LamarqueNo ratings yet
- Wang 2013Document7 pagesWang 2013Archisman MahapatraNo ratings yet
- Ascorbic Acid PaperDocument9 pagesAscorbic Acid PaperLaura Martinez RachadellNo ratings yet
- Dhami Et Al-2019-Journal of Neurochemistry PDFDocument18 pagesDhami Et Al-2019-Journal of Neurochemistry PDFJawad A. KhanNo ratings yet
- J. Biol. Chem.-1988-Meister-17205-8Document4 pagesJ. Biol. Chem.-1988-Meister-17205-8Pepé DubNo ratings yet
- Aquous Extract of Tamarind SeedsDocument11 pagesAquous Extract of Tamarind SeedsAmalia FirdhaNo ratings yet
- Proteosome Alteration Type2 Cell ARDSDocument15 pagesProteosome Alteration Type2 Cell ARDSelibb346No ratings yet
- Identifying Transglutaminase Reaction Products Via Mass Spectrometry As Exemplified by The MUC2 MucinDocument5 pagesIdentifying Transglutaminase Reaction Products Via Mass Spectrometry As Exemplified by The MUC2 MucinRosita HandayaniNo ratings yet
- A. 2015. Uzlikova, M. The Effect of MTZ On The Cell Cylce and DNA in MTZ-suscep. and Resistant. Giardia Cell LinesDocument7 pagesA. 2015. Uzlikova, M. The Effect of MTZ On The Cell Cylce and DNA in MTZ-suscep. and Resistant. Giardia Cell LinesJorge Luis RomeroNo ratings yet
- Mitochondria and Cell SignallingDocument9 pagesMitochondria and Cell SignallingNeto Pacífico NetoNo ratings yet
- Glutathione Centennial: Molecular Perspectives and Clinical ImplicationsFrom EverandGlutathione Centennial: Molecular Perspectives and Clinical ImplicationsNaoyuki TaniguchiRating: 5 out of 5 stars5/5 (1)
- Alatas 2016Document57 pagesAlatas 2016bagas umam alwiNo ratings yet
- ph0547s70Document20 pagesph0547s70bagas umam alwiNo ratings yet
- Moringaoleifera10 11648 J Ijnfs 20150406 14Document8 pagesMoringaoleifera10 11648 J Ijnfs 20150406 14bagas umam alwiNo ratings yet
- 2022 Article 231Document14 pages2022 Article 231bagas umam alwiNo ratings yet
- Jetr MS Id 555577Document4 pagesJetr MS Id 555577bagas umam alwiNo ratings yet
- 2020 Article 773Document10 pages2020 Article 773bagas umam alwiNo ratings yet
- Antidiabetic Drugs: Raymund N. Tapaoan, RPHDocument23 pagesAntidiabetic Drugs: Raymund N. Tapaoan, RPHNicole EncinaresNo ratings yet
- Vildagliptin: The Evidence For Its Place in The Treatment of Type 2 Diabetes MellitusDocument18 pagesVildagliptin: The Evidence For Its Place in The Treatment of Type 2 Diabetes Mellitusp k agarwalNo ratings yet
- Diabetes Review LiteratureDocument72 pagesDiabetes Review LiteratureAntonio Zandre Moreira LopesNo ratings yet
- Case 16 QuestionsDocument10 pagesCase 16 Questionsapi-532124328No ratings yet
- Pancreas: Anatomy, Diseases and Health Implications: September 2012Document53 pagesPancreas: Anatomy, Diseases and Health Implications: September 2012DrFarah Emad AliNo ratings yet
- Amaryl Prospect in EnglezaDocument14 pagesAmaryl Prospect in EnglezaLaura ZahariaNo ratings yet
- Ebook Endocrine Secrets 2 Full Chapter PDFDocument64 pagesEbook Endocrine Secrets 2 Full Chapter PDFjason.green261100% (24)
- Pathophysiology of Gestational Diabetes Mellitus: AuthorDocument10 pagesPathophysiology of Gestational Diabetes Mellitus: AuthorMayra PereiraNo ratings yet
- Medicinal Plants and Diabetes PDFDocument8 pagesMedicinal Plants and Diabetes PDFTowhid HasanNo ratings yet
- UG PROJECT ADocument56 pagesUG PROJECT AAdah EneNo ratings yet
- Galvus Met TabDocument23 pagesGalvus Met TabMaria Nicole EconasNo ratings yet
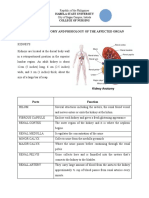
- Review of The Anatomy and Phisiology of The Affected Organ: Isabela State University College of NursingDocument7 pagesReview of The Anatomy and Phisiology of The Affected Organ: Isabela State University College of NursingWinnie AriolaNo ratings yet
- 8 Summary Major Chronic Illness of Older Adult Understanding The Pathophysiology and Nursing InterventionDocument21 pages8 Summary Major Chronic Illness of Older Adult Understanding The Pathophysiology and Nursing InterventionSteffiNo ratings yet
- Research ArticleDocument12 pagesResearch ArticleDivya RasoiNo ratings yet
- THE-EFFECTIVENESS-AND-EFFICIENCY-OF-SARSALIDA-LEAF (Repaired) - 033503Document32 pagesTHE-EFFECTIVENESS-AND-EFFICIENCY-OF-SARSALIDA-LEAF (Repaired) - 033503Bianca AndayaNo ratings yet
- Practical Guide To Insulin Therapy in Type 2 Diabetes: January 2011Document91 pagesPractical Guide To Insulin Therapy in Type 2 Diabetes: January 2011leeyie22No ratings yet
- Manuskrip TerjemahanDocument5 pagesManuskrip TerjemahanPika R97No ratings yet
- DSL: Diabetes Mellitus: Endocrine & Metabolism ModuleDocument45 pagesDSL: Diabetes Mellitus: Endocrine & Metabolism ModuleTower AlangNo ratings yet
- Diabetes - WMDocument15 pagesDiabetes - WMEmon KhanNo ratings yet
- 7 Answers To End-Of-Chapter QuestionsDocument6 pages7 Answers To End-Of-Chapter Questionsalyss ramdathNo ratings yet
- Neonatal Diabetes Mellitus: DR Nahim DM Neo Resident, ICH & HCDocument62 pagesNeonatal Diabetes Mellitus: DR Nahim DM Neo Resident, ICH & HCBhupendra GuptaNo ratings yet
- Type 1 Diabetes: Basis of Causes and Away of PreventionDocument11 pagesType 1 Diabetes: Basis of Causes and Away of PreventionOpenaccess Research paperNo ratings yet
- Pharmacotherapy of Diabetes MellitusDocument120 pagesPharmacotherapy of Diabetes MellitusElfiaNeswitaNo ratings yet
- Physiology DiabetesDocument48 pagesPhysiology Diabetesrajesh g100% (1)
- Case Study 22 Diabetes I KNH 411Document18 pagesCase Study 22 Diabetes I KNH 411api-30189722067% (3)
- Ben Lynch Blood Sugar GenesDocument37 pagesBen Lynch Blood Sugar GenesJazmin L. SolizNo ratings yet