You might also like
- Successes and Challenges of NK Immunotherapy: Breaking Tolerance to Cancer ResistanceFrom EverandSuccesses and Challenges of NK Immunotherapy: Breaking Tolerance to Cancer ResistanceNo ratings yet
- Fiedler 2016Document25 pagesFiedler 2016Puneet ChaudharyNo ratings yet
- Mrna 2Document12 pagesMrna 2api-675909478No ratings yet
- mRNA Lipid Nanoparticle-Mediated Pyr-Optosis Sensitizes Immunologically Cold Tumors To Checkpoint ImmunotherapyDocument16 pagesmRNA Lipid Nanoparticle-Mediated Pyr-Optosis Sensitizes Immunologically Cold Tumors To Checkpoint Immunotherapy0728lrhNo ratings yet
- Paper 3 - Immunomodulating Nano-Adaptors Potentiate Antibody-Based Cancer ImmunotherapyDocument14 pagesPaper 3 - Immunomodulating Nano-Adaptors Potentiate Antibody-Based Cancer ImmunotherapyChauPhuongNo ratings yet
- Science AbmDocument7 pagesScience AbmCosti ValiNo ratings yet
- Kinetics of mRNA Delivery and Protein TranslationDocument20 pagesKinetics of mRNA Delivery and Protein TranslationAkbar JuliansyahNo ratings yet
- Immune cell metabolism and metabolic reprogrammingDocument13 pagesImmune cell metabolism and metabolic reprogrammingFlávia PampolhaNo ratings yet
- Peptide-Mediated Delivery of CRISPR Enzymes For The Efficient Editing of Primary Human LymphocytesDocument17 pagesPeptide-Mediated Delivery of CRISPR Enzymes For The Efficient Editing of Primary Human LymphocytesBîkes NâçarNo ratings yet
- JC 18Document8 pagesJC 18emiliaclyde39No ratings yet
- mRNA VaccinesDocument27 pagesmRNA VaccinesBOMMIDI JAHNAVI (RA2132001010057)100% (1)
- Research Epstein-Barr virus encoded latent membraneDocument9 pagesResearch Epstein-Barr virus encoded latent membraneMiss AmyNo ratings yet
- Self Assembled Polymeric Micelle As A Novel MRNA 2021 Journal of ControlledDocument11 pagesSelf Assembled Polymeric Micelle As A Novel MRNA 2021 Journal of Controlledirene cayaNo ratings yet
- Crozza 2020-BioRxiv-ENPP1 Inhibitors - ReviewDocument37 pagesCrozza 2020-BioRxiv-ENPP1 Inhibitors - ReviewBrahmam PujalaNo ratings yet
- Assignment 7 noncoding small RNAsDocument3 pagesAssignment 7 noncoding small RNAsThảo Vy Nguyễn VươngNo ratings yet
- Stmcls 32 6 1553Document12 pagesStmcls 32 6 1553ThiKem NguyenNo ratings yet
- Recent Advances in mRNA Vaccine Delivery: Nano ResearchDocument17 pagesRecent Advances in mRNA Vaccine Delivery: Nano ResearchMETRY IMANDA PUTRINo ratings yet
- Huang Oncolytic Adeno Progr by Sythetic Gene Nature Com 2019Document15 pagesHuang Oncolytic Adeno Progr by Sythetic Gene Nature Com 2019Kudelko MatNo ratings yet
- Exp. Derm.-2008-Di Como-405-11Document7 pagesExp. Derm.-2008-Di Como-405-11Charles J. DiComo, PhDNo ratings yet
- Science Ade6967Document9 pagesScience Ade69670728lrhNo ratings yet
- Halloran 2004Document15 pagesHalloran 2004Agustina ParnisariNo ratings yet
- The CAMP Pathway As Therapeutic Target in Autoimmune and Inflammatory Diseases - PMCDocument21 pagesThe CAMP Pathway As Therapeutic Target in Autoimmune and Inflammatory Diseases - PMCRovin RamphalNo ratings yet
- Research-Highlights 2018 YmtheDocument2 pagesResearch-Highlights 2018 Ymtheedwin tjandraNo ratings yet
- ART. ScienceDocument20 pagesART. ScienceNICOLE ANDREA PICO TAMAYONo ratings yet
- p53-Regulated lncRNA Activates Enhancer DomainsDocument12 pagesp53-Regulated lncRNA Activates Enhancer DomainskNo ratings yet
- DNA Vaccines 2005Document7 pagesDNA Vaccines 2005Efrén ChávezNo ratings yet
- Dong Et Al 2022 - Nanoparticles (NPS) - Mediated Systemic mRNA Delivery To Reverse Trastuzumab Resistance For Effective Breast Cancer TherapyDocument12 pagesDong Et Al 2022 - Nanoparticles (NPS) - Mediated Systemic mRNA Delivery To Reverse Trastuzumab Resistance For Effective Breast Cancer TherapySazid AhmadNo ratings yet
- 2021_Schoenmaker_mRNA-lipid_nanoparticle_COVID-19_vaccines--Structure and stabilityDocument13 pages2021_Schoenmaker_mRNA-lipid_nanoparticle_COVID-19_vaccines--Structure and stabilitylienhartviktorNo ratings yet
- Systemic Biodistribution Expression Mice: Gene Therapy: and Long-Term of A Transgene inDocument5 pagesSystemic Biodistribution Expression Mice: Gene Therapy: and Long-Term of A Transgene inCa Nd RaNo ratings yet
- The Immune Microenvironment, Genome-Wide Copy Number Aberrations, and Survival in MesotheliomaDocument10 pagesThe Immune Microenvironment, Genome-Wide Copy Number Aberrations, and Survival in MesotheliomaTaloipaNo ratings yet
- CD28-dependent Rac1 Activation Is The Molecular Target of Azathioprine in Primary Human CD4 T LymphocytesDocument14 pagesCD28-dependent Rac1 Activation Is The Molecular Target of Azathioprine in Primary Human CD4 T LymphocytesasmanNo ratings yet
- MEK Inhibitors Activate WNT Signalling and Induce Stem Cell Plasticity in Colorectal CancerDocument17 pagesMEK Inhibitors Activate WNT Signalling and Induce Stem Cell Plasticity in Colorectal CancerYuqing LiangNo ratings yet
- Constitutive Transduction of Peptide Transporter and HLA Genes Restores Antigen ProcessingDocument6 pagesConstitutive Transduction of Peptide Transporter and HLA Genes Restores Antigen Processingroyhiranmay94No ratings yet
- GT 201646Document9 pagesGT 201646prima yogiNo ratings yet
- Lee 2012Document11 pagesLee 2012JA VelascoNo ratings yet
- Project ProposalDocument8 pagesProject ProposalArnab ChakrabortyNo ratings yet
- The Potential of Conditionally Replicating AdenovirusDocument1 pageThe Potential of Conditionally Replicating AdenovirusJairo BermúdezNo ratings yet
- Enhancing CRISPR/Cas Gene Editing Through Modulating Cellular Mechanical Properties For Cancer TherapyDocument16 pagesEnhancing CRISPR/Cas Gene Editing Through Modulating Cellular Mechanical Properties For Cancer TherapyCarolinaNo ratings yet
- Avalia 3Document23 pagesAvalia 3Marcus MvmNo ratings yet
- Comparing Immobilized Kinase Inhibitors and Covalent ATP Probes For Proteomic Profiling of Kinase Expression and Drug Selectivity.Document9 pagesComparing Immobilized Kinase Inhibitors and Covalent ATP Probes For Proteomic Profiling of Kinase Expression and Drug Selectivity.Darkill JackNo ratings yet
- Dandritic Cells ActivationDocument5 pagesDandritic Cells ActivationMazhar HussainNo ratings yet
- A Chemical Biology Screen Identifies Glucocorticoids That Regulate C - 2007 - BLDocument8 pagesA Chemical Biology Screen Identifies Glucocorticoids That Regulate C - 2007 - BLTareeqanwar MohammedNo ratings yet
- Tanshinone IIA Harmonizes The Crosstalk of Autophagy and 2019 InternationalDocument12 pagesTanshinone IIA Harmonizes The Crosstalk of Autophagy and 2019 InternationalfirdaNo ratings yet
- Lipid Nanoparticles-Loaded With Toxin mRNA Represents A New Strategy For The Treatment of Solid TumorsDocument12 pagesLipid Nanoparticles-Loaded With Toxin mRNA Represents A New Strategy For The Treatment of Solid Tumorshentaiaddict124No ratings yet
- Systemic mRNA Delivery To The Lungs by Functional Polyester-Based CarriersDocument25 pagesSystemic mRNA Delivery To The Lungs by Functional Polyester-Based CarriersRajat DasNo ratings yet
- 2000-Drug Development ResearchDocument10 pages2000-Drug Development ResearchEmerson CasaliNo ratings yet
- MMP10 NSCLCDocument9 pagesMMP10 NSCLCbhaskarNo ratings yet
- Please Use Endnote Program To Organize Your ReferencesDocument25 pagesPlease Use Endnote Program To Organize Your ReferencesMy PlanNo ratings yet
- Author Proof: Transmabs: Cell-Penetrating Antibodies, The Next GenerationDocument5 pagesAuthor Proof: Transmabs: Cell-Penetrating Antibodies, The Next GenerationVera MirandaNo ratings yet
- mRNA As A Transformative Technology For VaccineDocument6 pagesmRNA As A Transformative Technology For VaccineKlinik Hidayatullah MalangNo ratings yet
- Inmunoterapia en Cancer de PulmonDocument7 pagesInmunoterapia en Cancer de PulmonJorge ZegarraNo ratings yet
- Delivering The Messenger: Advances in Technologies For Therapeutic mRNA DeliveryDocument19 pagesDelivering The Messenger: Advances in Technologies For Therapeutic mRNA DeliveryMurilo Davoli FerreiraNo ratings yet
- 1995 JicrDocument9 pages1995 JicrAndrea la SalaNo ratings yet
- Nf-Κb-Modulated Mir-130A Targets Tnf-Α In Cervical Cancer CellsDocument14 pagesNf-Κb-Modulated Mir-130A Targets Tnf-Α In Cervical Cancer CellsMazhar HussainNo ratings yet
- Arn de InterferenciaDocument8 pagesArn de InterferenciaRosita Aguirre FloresNo ratings yet
- (Kyung Hee, Et Al, 2006) PGDocument4 pages(Kyung Hee, Et Al, 2006) PGWinda AlpiniawatiNo ratings yet
- Direct identification of clinically relevant neoepitopes presented on native human melanoma tissue by mass spectrometryDocument16 pagesDirect identification of clinically relevant neoepitopes presented on native human melanoma tissue by mass spectrometryZUNENo ratings yet
- 18-2021-Delorme-ANXA1 - PDMI33948376Document17 pages18-2021-Delorme-ANXA1 - PDMI33948376mercedesNo ratings yet
- Engineering Microbial Cell Morphology and Membranehomeostasis Toward Industrial ApplicationsDocument10 pagesEngineering Microbial Cell Morphology and Membranehomeostasis Toward Industrial ApplicationsElinaNo ratings yet
- Cisplatin-Resistant A549 Non-Small Cell Lung Cancer Cells Can Be Identified by Increased Mitochondrial Mass and Are Sensitive To Pemetrexed TreatmentDocument14 pagesCisplatin-Resistant A549 Non-Small Cell Lung Cancer Cells Can Be Identified by Increased Mitochondrial Mass and Are Sensitive To Pemetrexed Treatment高燕云No ratings yet
- HE - Food and Beverages CGDocument14 pagesHE - Food and Beverages CGJohn Leonard88% (8)
- Organozinc CompoundDocument8 pagesOrganozinc Compoundamanuel tafeseNo ratings yet
- TDS - Rheofinish 288 FDDocument2 pagesTDS - Rheofinish 288 FDVenkata RaoNo ratings yet
- Pro XP Manual Electrostatic Guns For Waterborne ApplicationsDocument4 pagesPro XP Manual Electrostatic Guns For Waterborne ApplicationsDiogo MatosNo ratings yet
- Surgery Hazel Final1Document21 pagesSurgery Hazel Final1Sittie RamosNo ratings yet
- Father Dámaso: María Clara Quotes in Noli Me TangereDocument11 pagesFather Dámaso: María Clara Quotes in Noli Me TangereSilver ArgentNo ratings yet
- St. Luke's Echocardiogram Patient GuideDocument2 pagesSt. Luke's Echocardiogram Patient Guideteju13aNo ratings yet
- DH 201 To 300 Bedded Revised Draft PDFDocument158 pagesDH 201 To 300 Bedded Revised Draft PDFAmit Kumar MishraNo ratings yet
- Observation of Unknown SpectrumDocument3 pagesObservation of Unknown SpectrumOmar HusseinNo ratings yet
- CA Prostate by Dr. Musaib MushtaqDocument71 pagesCA Prostate by Dr. Musaib MushtaqDr. Musaib MushtaqNo ratings yet
- Soal Bahasa Inggris Kelas Xi Semester 2 Petunjuk Khusus:: I Just Want To Be AloneDocument9 pagesSoal Bahasa Inggris Kelas Xi Semester 2 Petunjuk Khusus:: I Just Want To Be Alonesmk bhaktiasihNo ratings yet
- 06 CET Power - User Manual - Monitoring T1S T2S CanDis (X Web) - V1.0Document78 pages06 CET Power - User Manual - Monitoring T1S T2S CanDis (X Web) - V1.0Alfonso chaconNo ratings yet
- KOLHAN UNIVERSITY B.Sc Zoology Semester I Exam 2021 Provisional Marks CardDocument2 pagesKOLHAN UNIVERSITY B.Sc Zoology Semester I Exam 2021 Provisional Marks CardSmritiNo ratings yet
- How To Make A Literature Review TableDocument8 pagesHow To Make A Literature Review Tabletwfmadsif100% (1)
- BASF Puristar R3-12 - BF-9220 - PuriStar - R3-12 - PDS - Rev.2020-12 Spec SheetDocument2 pagesBASF Puristar R3-12 - BF-9220 - PuriStar - R3-12 - PDS - Rev.2020-12 Spec SheetAlNo ratings yet
- Adime Malnutrition and OncolgyDocument6 pagesAdime Malnutrition and Oncolgyapi-300587226100% (1)
- HDFC Life InsuranceDocument12 pagesHDFC Life Insurancesaswat mohantyNo ratings yet
- VolvoTD70 Service Manual EngineDocument110 pagesVolvoTD70 Service Manual EngineHeikki Alakontiola87% (47)
- Refrigeration and Air Conditioning Technology 8th Edition Tomczyk Solutions ManualDocument36 pagesRefrigeration and Air Conditioning Technology 8th Edition Tomczyk Solutions Manualeelspearbetrayal.yagz100% (52)
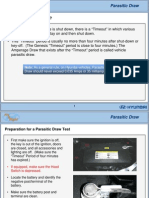
- Parasitic Draw ChartDocument9 pagesParasitic Draw Chartlionellin83No ratings yet
- Calcium chloride MSDSDocument5 pagesCalcium chloride MSDSDarshilNo ratings yet
- Measuring Blood Pressure AccuratelyDocument2 pagesMeasuring Blood Pressure AccuratelyxmantestNo ratings yet
- August 2016Document15 pagesAugust 2016Pumper Trader0% (1)
- Wisdom Chi KungDocument0 pagesWisdom Chi KungDevlinPyxNo ratings yet
- Gym Membership AgreementDocument3 pagesGym Membership Agreementzde thai100% (1)
- Diagnostic Test Science 5Document7 pagesDiagnostic Test Science 5Rex Russel SalemNo ratings yet
- Fuel CellDocument71 pagesFuel CellAhmed Mostafa100% (1)
- PDF Fcuk CTLG 2020 Split 01Document46 pagesPDF Fcuk CTLG 2020 Split 01Ted Andrew AbalosNo ratings yet
- CyberstalkingDocument22 pagesCyberstalkingManasa M DharNo ratings yet
- ETU 776 TripDocument1 pageETU 776 TripbhaskarinvuNo ratings yet
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (402)
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (15)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeNo ratings yet
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 5 out of 5 stars5/5 (4)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (169)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (78)
- The Happiness Trap: How to Stop Struggling and Start LivingFrom EverandThe Happiness Trap: How to Stop Struggling and Start LivingRating: 4 out of 5 stars4/5 (1)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (42)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- Techniques Exercises And Tricks For Memory ImprovementFrom EverandTechniques Exercises And Tricks For Memory ImprovementRating: 4.5 out of 5 stars4.5/5 (40)
- The Ultimate Guide To Memory Improvement TechniquesFrom EverandThe Ultimate Guide To Memory Improvement TechniquesRating: 5 out of 5 stars5/5 (34)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingFrom EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingRating: 3.5 out of 5 stars3.5/5 (33)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisFrom EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (1)
- Summary: It Didn't Start with You: How Inherited Family Trauma Shapes Who We Are and How to End the Cycle By Mark Wolynn: Key Takeaways, Summary & AnalysisFrom EverandSummary: It Didn't Start with You: How Inherited Family Trauma Shapes Who We Are and How to End the Cycle By Mark Wolynn: Key Takeaways, Summary & AnalysisRating: 5 out of 5 stars5/5 (3)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryFrom EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryRating: 4 out of 5 stars4/5 (44)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaFrom EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaRating: 4.5 out of 5 stars4.5/5 (266)
- The Tennis Partner: A Doctor's Story of Friendship and LossFrom EverandThe Tennis Partner: A Doctor's Story of Friendship and LossRating: 4.5 out of 5 stars4.5/5 (4)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (328)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingFrom EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingRating: 5 out of 5 stars5/5 (5)