You might also like
- Biofumigation: A Potential Aspect For Suppression of Plant-Parasitic NematodesDocument7 pagesBiofumigation: A Potential Aspect For Suppression of Plant-Parasitic NematodesIJEAB JournalNo ratings yet
- tmpD493 TMPDocument11 pagestmpD493 TMPFrontiersNo ratings yet
- Bashan y Olguin 1998Document4 pagesBashan y Olguin 1998María Flavia LunaNo ratings yet
- Plant Growth Promoting Rhizobacteria and Sustainable Agriculture: A ReviewDocument6 pagesPlant Growth Promoting Rhizobacteria and Sustainable Agriculture: A ReviewMohammad Mosharraf HossainNo ratings yet
- PGPRs ReviewDocument15 pagesPGPRs ReviewMuhammad Asif RoyNo ratings yet
- Seed Biopriming PGPR IntrodDocument14 pagesSeed Biopriming PGPR IntrodMohammad Mosharraf HossainNo ratings yet
- Trichoderma Species: Versatile Plant SymbiontsDocument11 pagesTrichoderma Species: Versatile Plant SymbiontsAdam StankiewiczNo ratings yet
- Their Potential PGPR ReviewDocument19 pagesTheir Potential PGPR ReviewMohammad Mosharraf HossainNo ratings yet
- Diversity of Plant Growth Promoting Rhizobacteria Associated With MaizeDocument23 pagesDiversity of Plant Growth Promoting Rhizobacteria Associated With MaizeoigresvaxNo ratings yet
- Azospirillum Spp. in Grasses and Forages. Review: AbstractDocument18 pagesAzospirillum Spp. in Grasses and Forages. Review: AbstractSandro ManchegoNo ratings yet
- Chapter 25: Azospirillum Spp. and Related Pgprs Inocula Use in Intensive AgricultureDocument18 pagesChapter 25: Azospirillum Spp. and Related Pgprs Inocula Use in Intensive AgricultureGabyFasciglioneNo ratings yet
- Beneficial Bacteria of Agricultural ImportanceDocument12 pagesBeneficial Bacteria of Agricultural ImportanceXevin MoreiraNo ratings yet
- Soil Beneficial Bacteria and Their Role in Plant GrowthDocument20 pagesSoil Beneficial Bacteria and Their Role in Plant GrowthKhairilanuar Mohd HanimNo ratings yet
- Fsufs 05 606815Document23 pagesFsufs 05 606815antonioNo ratings yet
- International Journal of Food Microbiology 334 (2020) 108799Document11 pagesInternational Journal of Food Microbiology 334 (2020) 108799Tuğçe KanıkNo ratings yet
- 10glyphosate Effects On Soil Rhizosphere-Associated Bacterial CommunitiesDocument6 pages10glyphosate Effects On Soil Rhizosphere-Associated Bacterial CommunitiesJOSUENo ratings yet
- Endophytic Fungi in Economically Important Plants: Ecological Aspects, Diversity and Potential Biotechnological ApplicationsDocument14 pagesEndophytic Fungi in Economically Important Plants: Ecological Aspects, Diversity and Potential Biotechnological ApplicationsFabian Giovanny Márquez NiñoNo ratings yet
- Contreras Cornejo 2017Document9 pagesContreras Cornejo 2017Karen Avalos VelaNo ratings yet
- 1 s2.0 S1161030121000654 MainDocument12 pages1 s2.0 S1161030121000654 MainAB LimaNo ratings yet
- Brassinosteroid Analogues Effect On Yield and Quality Parameters of Field-Grown Lettuce (Lactuca Sativa L.)Document9 pagesBrassinosteroid Analogues Effect On Yield and Quality Parameters of Field-Grown Lettuce (Lactuca Sativa L.)lucasgirioNo ratings yet
- Percobaan Hidrolisat Mikroalga Dengan Aplikasi Foliar Pada Pertumbuhan Petunia X HybridaDocument7 pagesPercobaan Hidrolisat Mikroalga Dengan Aplikasi Foliar Pada Pertumbuhan Petunia X Hybridabayu garinNo ratings yet
- Agrultura MicrobiosDocument25 pagesAgrultura Microbioschobyts27lizyNo ratings yet
- Backer 2018Document17 pagesBacker 2018adham ahmedNo ratings yet
- 1 s2.0 S0167880916304820 MainDocument9 pages1 s2.0 S0167880916304820 MainQuevedodi IqdNo ratings yet
- Quorum Sensing and Indole-3-Acetic Acid Degradation Play A Role in Colonization and Plant Growth Promotion of Arabidopsis Thaliana by Burkholderia Phytofirmans PSJNDocument9 pagesQuorum Sensing and Indole-3-Acetic Acid Degradation Play A Role in Colonization and Plant Growth Promotion of Arabidopsis Thaliana by Burkholderia Phytofirmans PSJNFrancisca MartinichNo ratings yet
- 2020 Effects of Plant Growth Promoting MicroorganismsDocument19 pages2020 Effects of Plant Growth Promoting MicroorganismsFabian Giovanny Márquez NiñoNo ratings yet
- 1 s2.0 S094450131300116X MainDocument7 pages1 s2.0 S094450131300116X MainJorge CorderoNo ratings yet
- Article 1 Biofertilizers and Sustainable AgricultureDocument12 pagesArticle 1 Biofertilizers and Sustainable AgriculturePaolita CortesNo ratings yet
- Biosynthesis and Function of Terpenoid Defense Compounds in Maize (Zea Mays)Document10 pagesBiosynthesis and Function of Terpenoid Defense Compounds in Maize (Zea Mays)Silvio FaveroNo ratings yet
- Biofumigation: An Alternative Strategy For The Control of Plant Parasitic NematodesDocument11 pagesBiofumigation: An Alternative Strategy For The Control of Plant Parasitic Nematodestehmina sharifNo ratings yet
- Daval Et Al., 2020 - Soil Microbiota Influences Clubroot DiseaseDocument25 pagesDaval Et Al., 2020 - Soil Microbiota Influences Clubroot DiseaseJorge CorderoNo ratings yet
- Environmental Pollution: Manli Duan, Haichao Li, Jie Gu, Xiaxia Tuo, Wei Sun, Xun Qian, Xiaojuan WangDocument9 pagesEnvironmental Pollution: Manli Duan, Haichao Li, Jie Gu, Xiaxia Tuo, Wei Sun, Xun Qian, Xiaojuan WangGeorgiana-LuizaNo ratings yet
- Effect Glifosato of Metabolism Stres and Nutritional Condition Bullfrog 2019Document15 pagesEffect Glifosato of Metabolism Stres and Nutritional Condition Bullfrog 2019JUAN CARLOS LGNo ratings yet
- Biocatalysis and Agricultural BiotechnologyDocument7 pagesBiocatalysis and Agricultural BiotechnologyLydiaNo ratings yet
- Nitrogen Is One of The Essential Elements Required For The Synthesis of Amino Acids WhichDocument11 pagesNitrogen Is One of The Essential Elements Required For The Synthesis of Amino Acids Whichsanjay kumarNo ratings yet
- editors,+Journal+editor,+JANS+7 (1) 540-556 (2015)Document17 pageseditors,+Journal+editor,+JANS+7 (1) 540-556 (2015)544bvxfffsNo ratings yet
- Everything You Must Know About Azospirillum and Its Impact On Agriculture and BeyondDocument19 pagesEverything You Must Know About Azospirillum and Its Impact On Agriculture and BeyondsausanNo ratings yet
- Unlocking The Potential of Plant Growth-Promoting Rhizobacteria On Soil Health and The Sustainability of Agricultural SystemsDocument20 pagesUnlocking The Potential of Plant Growth-Promoting Rhizobacteria On Soil Health and The Sustainability of Agricultural Systemsmazhar.commereceNo ratings yet
- 1415 4757 GMB S1415 475738420150053Document19 pages1415 4757 GMB S1415 475738420150053this hihiNo ratings yet
- The Comparative Study of Biofertilizers On Growth and Yield of Wheat: A ReviewDocument6 pagesThe Comparative Study of Biofertilizers On Growth and Yield of Wheat: A ReviewInternational Journal of Innovative Science and Research Technology100% (1)
- Cisgenesis and Intragenesis: New Tools For Improving CropsDocument10 pagesCisgenesis and Intragenesis: New Tools For Improving CropsgeraldineChrisNo ratings yet
- JTPR 02Document15 pagesJTPR 02kasahun AmareNo ratings yet
- Impact of Antifungals Producing Rhizobacteria On The Performance of Vigna Radiata in The Presence of Arbuscular Mycorrhizal FungiDocument12 pagesImpact of Antifungals Producing Rhizobacteria On The Performance of Vigna Radiata in The Presence of Arbuscular Mycorrhizal FungimetbijayNo ratings yet
- Gluconacetobacter Diazotrophicus: An Overview: February 2014Document11 pagesGluconacetobacter Diazotrophicus: An Overview: February 2014crisNo ratings yet
- Bacterial Endophytes: Recent Developments and ApplicationsDocument9 pagesBacterial Endophytes: Recent Developments and ApplicationsmaczweiNo ratings yet
- ENGENVI Case StudyDocument8 pagesENGENVI Case StudyThrowaway TwoNo ratings yet
- Sachin44 50Document7 pagesSachin44 50MogleNo ratings yet
- Degradation Features of Pesticides A Review On (Metallo) Porphyrin-Mediated Catalytic ProcessesDocument20 pagesDegradation Features of Pesticides A Review On (Metallo) Porphyrin-Mediated Catalytic ProcessesIasmin ResendeNo ratings yet
- Art:10.1007/s11101 006 9032 2Document35 pagesArt:10.1007/s11101 006 9032 2Caroline LopesNo ratings yet
- The Potentiality of Selected PGPR - Azotobacter, For Sustainable Agriculture in IndiaDocument5 pagesThe Potentiality of Selected PGPR - Azotobacter, For Sustainable Agriculture in IndiaSyarafina Ratna PutriNo ratings yet
- Phytochemistry: Nicol As Cara, Patricia N. Piccoli, Leonardo Bolcato, Carlos F. Marfil, Ricardo W. MasuelliDocument8 pagesPhytochemistry: Nicol As Cara, Patricia N. Piccoli, Leonardo Bolcato, Carlos F. Marfil, Ricardo W. MasuelliLuisNo ratings yet
- Bacterial Siderophoreasa Plant Growth PROMOTERDocument18 pagesBacterial Siderophoreasa Plant Growth PROMOTEREdgar Meneses MoranNo ratings yet
- Arroz Temperaturas Nocturnas IIDocument7 pagesArroz Temperaturas Nocturnas IIJorge Andres Jola HernandezNo ratings yet
- Plant Viral Disease ManagementDocument15 pagesPlant Viral Disease ManagementGadusoNo ratings yet
- Ajilogba-Babalola2019 Article GCMSAnalysisOfVolatileOrganicCDocument19 pagesAjilogba-Babalola2019 Article GCMSAnalysisOfVolatileOrganicCJulli StefanyNo ratings yet
- Aab 12420 PDFDocument14 pagesAab 12420 PDFEvelin CARLIERNo ratings yet
- Enhancement of Phenolics Flavonoids and Glucosinolates of Broccoli Brassica Olaracea Var ItalicaDocument8 pagesEnhancement of Phenolics Flavonoids and Glucosinolates of Broccoli Brassica Olaracea Var ItalicaNo VivoNo ratings yet
- Plant Growth Promoting Rhizobacteria (PGPR) : A Review: JUNE 2015Document13 pagesPlant Growth Promoting Rhizobacteria (PGPR) : A Review: JUNE 2015ViníciusNo ratings yet
- Classification and Naming of Plants: ExtensionDocument8 pagesClassification and Naming of Plants: ExtensionOrmie ChanNo ratings yet
- Dry Matter Production and Root System Development of Rice Cultivars Under Fluctuating Soil MoistureDocument12 pagesDry Matter Production and Root System Development of Rice Cultivars Under Fluctuating Soil MoistureErika MatiasNo ratings yet
- Annals of Agricultural Sciences: Nur A. Zin, Noor A. BadaluddinDocument11 pagesAnnals of Agricultural Sciences: Nur A. Zin, Noor A. BadaluddinAde regaNo ratings yet
- PS and CRDocument34 pagesPS and CRErika MatiasNo ratings yet
- Lab Report Plant TissuesDocument8 pagesLab Report Plant TissuesMOHD MU'IZZ BIN MOHD SHUKRI100% (6)
- Effects of Alternate Wetting and Drying Technique On Greenhouse Gas Emissions From Irrigated Rice Paddy in Central Luzon, PhilippinesDocument9 pagesEffects of Alternate Wetting and Drying Technique On Greenhouse Gas Emissions From Irrigated Rice Paddy in Central Luzon, PhilippinesErika MatiasNo ratings yet
- 2013 USSIRI - Soil Emission of Nitrous Oxide and Its MitigationDocument390 pages2013 USSIRI - Soil Emission of Nitrous Oxide and Its MitigationAnderson Dias SilveiraNo ratings yet
- Structures of Amino AcidsDocument32 pagesStructures of Amino AcidsUsman GhaniNo ratings yet
- Group Presentation RubricDocument1 pageGroup Presentation RubricErika MatiasNo ratings yet
- Project #3: Three-Minute Vlog: Criteria/ Ratings 4 3 2 1Document1 pageProject #3: Three-Minute Vlog: Criteria/ Ratings 4 3 2 1Erika MatiasNo ratings yet
- Moles, Units, and Conversion FactorsDocument19 pagesMoles, Units, and Conversion FactorsJermyn G EvangelistaNo ratings yet
- Qualitative Test - Amino Acids and ProteinsDocument9 pagesQualitative Test - Amino Acids and Proteinstapans_8No ratings yet
- Group Presentation RubricDocument1 pageGroup Presentation RubricErika MatiasNo ratings yet
- ORGANELLE Table PDFDocument2 pagesORGANELLE Table PDFDebjit BanerjeeNo ratings yet
- Prereg Drug Calculations Roy Sinclair Booklet 1Document66 pagesPrereg Drug Calculations Roy Sinclair Booklet 1Sudhakar KuppireddyNo ratings yet
- Mammalian Digestive Tracts: Mouth: Esophagus: Stomach: Small Intestine: Caecum: Large Intestine: RectumDocument19 pagesMammalian Digestive Tracts: Mouth: Esophagus: Stomach: Small Intestine: Caecum: Large Intestine: RectumErika MatiasNo ratings yet
- The Integumentary Morphology of Modern Birds-An Overview: 168 Croydon Turnpike, Plainfield, New Hampshire 03781-5403Document17 pagesThe Integumentary Morphology of Modern Birds-An Overview: 168 Croydon Turnpike, Plainfield, New Hampshire 03781-5403Erika MatiasNo ratings yet
- Washington State Cat Project Unit 3Document44 pagesWashington State Cat Project Unit 3CatalinAlbuNo ratings yet
- How To Solve Drug Dosage ProblemsDocument9 pagesHow To Solve Drug Dosage Problemspatra fadmaNo ratings yet
- How Do Animals Breathe?: All Vertebrates Lungs DiaphragmDocument1 pageHow Do Animals Breathe?: All Vertebrates Lungs DiaphragmErika MatiasNo ratings yet
- Aves SkinDocument7 pagesAves SkinErika MatiasNo ratings yet
- The Nervous System 2Document84 pagesThe Nervous System 2Erika MatiasNo ratings yet
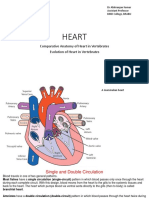
- Heart: Comparative Anatomy of Heart in Vertebrates Evolution of Heart in VertebratesDocument28 pagesHeart: Comparative Anatomy of Heart in Vertebrates Evolution of Heart in VertebratesErika MatiasNo ratings yet
- Heart: Comparative Anatomy of Heart in Vertebrates Evolution of Heart in VertebratesDocument28 pagesHeart: Comparative Anatomy of Heart in Vertebrates Evolution of Heart in VertebratesErika MatiasNo ratings yet
- Circulatory SystemDocument23 pagesCirculatory SystemErika MatiasNo ratings yet
- Comparative Anatomy of Heart in VertebratesDocument8 pagesComparative Anatomy of Heart in VertebratesErika MatiasNo ratings yet
- Comparative Anatomy of Heart in VertebratesDocument8 pagesComparative Anatomy of Heart in VertebratesErika MatiasNo ratings yet
- Mammalian Digestive Tracts: Mouth: Esophagus: Stomach: Small Intestine: Caecum: Large Intestine: RectumDocument19 pagesMammalian Digestive Tracts: Mouth: Esophagus: Stomach: Small Intestine: Caecum: Large Intestine: RectumErika MatiasNo ratings yet
- Circulatory SystemDocument23 pagesCirculatory SystemErika MatiasNo ratings yet
- Proline Protocol: Bates Et Al. (1973) : G Proline/ml X ML Toluene X G of SampleDocument2 pagesProline Protocol: Bates Et Al. (1973) : G Proline/ml X ML Toluene X G of SampleErika MatiasNo ratings yet
- Nutrisi Cedera KepalaDocument46 pagesNutrisi Cedera Kepalatsunami555No ratings yet
- PhysioEx Exercise 9 Activity 5Document3 pagesPhysioEx Exercise 9 Activity 5Gonzeilo11No ratings yet
- Mumbai AyurvedaDocument2 pagesMumbai AyurvedaAditya SinghNo ratings yet
- 1 Il-10 ElisaDocument15 pages1 Il-10 ElisaYousra ZeidanNo ratings yet
- TranslationDocument1 pageTranslationGianne AbcedeNo ratings yet
- Choice of Enterostoma: Feeding Jejunostomy IleostomyDocument4 pagesChoice of Enterostoma: Feeding Jejunostomy IleostomyFikranaya SalimNo ratings yet
- Small IntestineDocument28 pagesSmall IntestineHusnainAhmedNo ratings yet
- Sample Client Training Program PeterDocument10 pagesSample Client Training Program PeterRaj Nandwani0% (1)
- Ineffective Airway Clearance NCPDocument1 pageIneffective Airway Clearance NCPBenz ParCoNo ratings yet
- RH Typing ReagentsDocument2 pagesRH Typing ReagentsMelanie Tomita100% (1)
- Class XI Practicals Term2Document5 pagesClass XI Practicals Term2Prince KNo ratings yet
- Peripheral Nerve Repair and RegenerationDocument15 pagesPeripheral Nerve Repair and RegenerationjaimeaquilesrinconNo ratings yet
- 24 Cerebral Blood FlowDocument31 pages24 Cerebral Blood FlowZuhaib Ahmed100% (1)
- NCP 1Document1 pageNCP 1hsiriaNo ratings yet
- Chloride & Protein in CSF: Biochemistry Lab - 3 StageDocument19 pagesChloride & Protein in CSF: Biochemistry Lab - 3 Stagebknmy2nx9qNo ratings yet
- A Visit To A GrandmotherDocument3 pagesA Visit To A GrandmotherHEY ERLNo ratings yet
- Knight, Angela - The VampdomDocument8 pagesKnight, Angela - The VampdomLaviniaNo ratings yet
- Laboratory Test Report: Mrs. V Valli Kumari 81 Years FemaleDocument2 pagesLaboratory Test Report: Mrs. V Valli Kumari 81 Years FemaleumaNo ratings yet
- Immunoglobulins/Antibody: Presented By: Shashi Regd. No. 11006142 Roll No. B15 Section RP8003Document17 pagesImmunoglobulins/Antibody: Presented By: Shashi Regd. No. 11006142 Roll No. B15 Section RP8003Shashi SharmaNo ratings yet
- SCIENCE 9-Lesson 1 PhotosynthesisDocument2 pagesSCIENCE 9-Lesson 1 PhotosynthesisTeacher anaNo ratings yet
- Foundation of Nursing Practice Exam For Nursing Process Physical and Health Assessment and Routine ProceduresDocument24 pagesFoundation of Nursing Practice Exam For Nursing Process Physical and Health Assessment and Routine ProceduresAdrian Miguel100% (1)
- Dr. Vijetha Shenoy Belle Associate Professor & In-Charge Clinical Biochemistry Laboratory Department of Biochemistry KMC ManipalDocument53 pagesDr. Vijetha Shenoy Belle Associate Professor & In-Charge Clinical Biochemistry Laboratory Department of Biochemistry KMC ManipalKavya GaneshNo ratings yet
- Dr. Lakshmi Kiruba N Assistant Professor Department of Anatomy SmbtimsrcDocument25 pagesDr. Lakshmi Kiruba N Assistant Professor Department of Anatomy Smbtimsrclakshmi kirubaNo ratings yet
- Kreisberg-Zakarin Et Al IPNS Antonie Van Leeuwenhoek 1999Document8 pagesKreisberg-Zakarin Et Al IPNS Antonie Van Leeuwenhoek 1999ElNo ratings yet
- 6.1 Digestion BNDocument24 pages6.1 Digestion BNHana ElattarNo ratings yet
- Transfer To Definitive Care EcaDocument40 pagesTransfer To Definitive Care EcaFahrurido Kusbari100% (1)
- Fluid Management BurnDocument13 pagesFluid Management BurnWeronika StarrNo ratings yet
- Facial Nerve MCQDocument4 pagesFacial Nerve MCQIqbal Syed100% (1)
- Sorenson Atlas of Human Histology Chapters 1 and 14Document56 pagesSorenson Atlas of Human Histology Chapters 1 and 14Nick john CaminadeNo ratings yet
- Immunology Dental Caries PDFDocument29 pagesImmunology Dental Caries PDFTio AjhaNo ratings yet