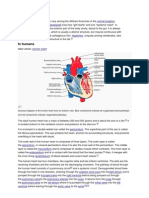
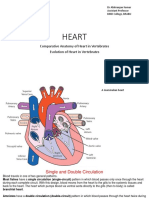
You might also like
- Lab Report Plant TissuesDocument8 pagesLab Report Plant TissuesMOHD MU'IZZ BIN MOHD SHUKRI100% (6)
- Structural Organisation in AnimalsDocument49 pagesStructural Organisation in AnimalsPukazhvanthen Paramanandhan100% (1)
- GEC08-SCIENCE, TECHNOLOGY & SOCIETY: Pandemics Past & PresentDocument1 pageGEC08-SCIENCE, TECHNOLOGY & SOCIETY: Pandemics Past & PresentErika MatiasNo ratings yet
- Prenatal and Postnatal Growth of the Cranial Vault, Base and MidfaceDocument219 pagesPrenatal and Postnatal Growth of the Cranial Vault, Base and MidfaceNizam Muhamad0% (1)
- Evolution of the Circulatory System: From Unicellular to MammalsDocument16 pagesEvolution of the Circulatory System: From Unicellular to MammalsSaajid AmraNo ratings yet
- Evolution of Heart in VertebratesDocument4 pagesEvolution of Heart in Vertebratesनितिन कुमारNo ratings yet
- B.Sc.I ZoologyDocument8 pagesB.Sc.I ZoologyGanesh YegadeNo ratings yet
- Dentition in Mammals CC-8 PDFDocument5 pagesDentition in Mammals CC-8 PDFtapas.manushNo ratings yet
- Evolution Origins TracedDocument96 pagesEvolution Origins TracedMendes ArmyNo ratings yet
- The Circulatory System - Lecture STEMDocument50 pagesThe Circulatory System - Lecture STEMMargeory Calatan100% (1)
- Vertebrate Respiratory Systems in 40 CharactersDocument129 pagesVertebrate Respiratory Systems in 40 Charactersgerman guazaNo ratings yet
- COMPARATIVE CIRCULATORY SYSTEMSDocument15 pagesCOMPARATIVE CIRCULATORY SYSTEMSSmartcool SoNo ratings yet
- Mutation: Neeraj Kumar Maurya Environmental BiotechnologyDocument24 pagesMutation: Neeraj Kumar Maurya Environmental BiotechnologyNeeraj MauryaNo ratings yet
- Ecology II Part B - Ecological Interactions - Food Chains Food Webs 2017Document38 pagesEcology II Part B - Ecological Interactions - Food Chains Food Webs 2017Shahriar HaqueNo ratings yet
- Amoeba PDFDocument5 pagesAmoeba PDFmanoj_rkl_07100% (1)
- Movement and Muscle MechanismsDocument12 pagesMovement and Muscle Mechanismsadithya4rajNo ratings yet
- India Quiz QuestionsDocument12 pagesIndia Quiz Questionsankur0708No ratings yet
- Animal KingdomDocument22 pagesAnimal KingdomdellfrogNo ratings yet
- Describe The Taxonomical of Identification of Cultivable FishesDocument5 pagesDescribe The Taxonomical of Identification of Cultivable FishesNarasimha MurthyNo ratings yet
- Respiratory SystemDocument138 pagesRespiratory SystemJosefantastic 75No ratings yet
- 4 Porifera Spicules & Canal SystemDocument34 pages4 Porifera Spicules & Canal Systemapi-3732735100% (2)
- Economic ZoologyDocument47 pagesEconomic ZoologyAntony SebastianNo ratings yet
- Archaeozoology: Camelopardalis Brisson, From The Prehistoric Site of Susunia, District BankuraDocument7 pagesArchaeozoology: Camelopardalis Brisson, From The Prehistoric Site of Susunia, District BankuraDibyendu DasNo ratings yet
- Human Digestive SystemDocument29 pagesHuman Digestive SystemRagini Dubey100% (1)
- Phylum PoriferaDocument30 pagesPhylum PoriferaEloisa Lopez100% (2)
- CamScanner Scans PDF DocsDocument6 pagesCamScanner Scans PDF DocsAdarsh jainNo ratings yet
- Comparative Vertebrate Anatomy Lecture Notes - Digestive SystemDocument14 pagesComparative Vertebrate Anatomy Lecture Notes - Digestive SystemMielah Ruth100% (1)
- Human Body Circulatory SystemDocument41 pagesHuman Body Circulatory SystemFrancis MacasioNo ratings yet
- Zoology B.sc.Document25 pagesZoology B.sc.sabiha yasmeenNo ratings yet
- Eukaryotic Genome Complexity - Learn Science at ScitableDocument4 pagesEukaryotic Genome Complexity - Learn Science at ScitableAnder ManaresNo ratings yet
- Wetlands of AssamDocument7 pagesWetlands of AssamHasibuddin AhmedNo ratings yet
- Practical XII TH STD Zoology EMDocument24 pagesPractical XII TH STD Zoology EMsanthoshsivanandam SivanandamNo ratings yet
- Ch. 7 (Homeostasis-Osmoregulation, Endocrine, Temperature)Document45 pagesCh. 7 (Homeostasis-Osmoregulation, Endocrine, Temperature)ipulNo ratings yet
- ZOOLOGY PRESENTATION: CONVICT CICHLIDDocument8 pagesZOOLOGY PRESENTATION: CONVICT CICHLIDਰੋਹਿਤ ਕੁਮਾਰNo ratings yet
- Canal System in Porifera: Ascon, Sycon and Leucon TypesDocument4 pagesCanal System in Porifera: Ascon, Sycon and Leucon TypesZotei RawihteNo ratings yet
- 0smoregulation in Fresh Water FishesDocument4 pages0smoregulation in Fresh Water FishesKazi AfrozNo ratings yet
- Abstracts For 32nd Congrass of ZoologyDocument350 pagesAbstracts For 32nd Congrass of ZoologySaima WaseemNo ratings yet
- Cnidaria SummaryDocument12 pagesCnidaria SummaryJose ArroyoNo ratings yet
- Most Common Edible Marine Fishes in KeralaDocument16 pagesMost Common Edible Marine Fishes in KeralaAneesh B LalNo ratings yet
- Lec4 SocialInsects TheSociety NotesDocument3 pagesLec4 SocialInsects TheSociety NotesAlexandra LvNo ratings yet
- Composite Fish Culture in HillsDocument20 pagesComposite Fish Culture in HillsSullip MajhiNo ratings yet
- Farooq Ali: Presented byDocument22 pagesFarooq Ali: Presented byFarooq AliNo ratings yet
- Treatment of Pediculosis Capitis - A Critical Appraisal of The Current LiteratureDocument12 pagesTreatment of Pediculosis Capitis - A Critical Appraisal of The Current Literaturejft842No ratings yet
- Oxidative Phosphorylation V Inhibitors and UncouplersDocument15 pagesOxidative Phosphorylation V Inhibitors and UncouplersIffatnazNo ratings yet
- NWIA National BrochureDocument32 pagesNWIA National BrochureSaurabh SinghviNo ratings yet
- OsmoregulationDocument14 pagesOsmoregulationmmills8No ratings yet
- FishesDocument74 pagesFishesLophyn UmarNo ratings yet
- Ento231 PDFDocument151 pagesEnto231 PDFMunna GuptaNo ratings yet
- TOrsionDocument19 pagesTOrsionDrSneha VermaNo ratings yet
- General Characteristic Features of Superclass PiscesDocument2 pagesGeneral Characteristic Features of Superclass PiscesSKNo ratings yet
- Common freshwater fishes of KeralaDocument18 pagesCommon freshwater fishes of KeralaVignesh Jayan100% (1)
- Transcription: RNA Polymerases and General Transcription FactorsDocument77 pagesTranscription: RNA Polymerases and General Transcription FactorsmluluNo ratings yet
- Systematic, Anatomy & Development PDFDocument1 pageSystematic, Anatomy & Development PDFAhmed RazaNo ratings yet
- Protochordata-Characters & PhylogenyDocument4 pagesProtochordata-Characters & PhylogenyAakash VNo ratings yet
- Roles and Impacts of Insects in AgricultureDocument58 pagesRoles and Impacts of Insects in Agriculturenovita manaluNo ratings yet
- Phylum Chordata Characteristics and ClassesDocument25 pagesPhylum Chordata Characteristics and ClassesMae Uayan Prisco-TagaraoNo ratings yet
- Most Common Edible Freshwater Fishes in KeralaDocument6 pagesMost Common Edible Freshwater Fishes in KeralaAneesh B LalNo ratings yet
- Chordate Characteristics and ClassificationDocument28 pagesChordate Characteristics and ClassificationNil BanerjeeNo ratings yet
- Origin and evolution theories of the chordate phylumDocument3 pagesOrigin and evolution theories of the chordate phylummonique100% (2)
- Respiratory SurfacesDocument11 pagesRespiratory SurfacesNitesh KotianNo ratings yet
- LearningPacket COMPACirculatorySystemDocument7 pagesLearningPacket COMPACirculatorySystemLei Jenevive UmbayNo ratings yet
- The Structure and Function of the Heart Across the Animal KingdomDocument9 pagesThe Structure and Function of the Heart Across the Animal KingdomDanagalla Venkat100% (2)
- Cardiovascular System - Heart: AssignmentDocument14 pagesCardiovascular System - Heart: AssignmentKriss MisraNo ratings yet
- PS and CRDocument34 pagesPS and CRErika MatiasNo ratings yet
- Dry Matter Production and Root System Development of Rice Cultivars Under Fluctuating Soil MoistureDocument12 pagesDry Matter Production and Root System Development of Rice Cultivars Under Fluctuating Soil MoistureErika MatiasNo ratings yet
- Classification and Naming of Plants: ExtensionDocument8 pagesClassification and Naming of Plants: ExtensionOrmie ChanNo ratings yet
- Annals of Agricultural Sciences: Nur A. Zin, Noor A. BadaluddinDocument11 pagesAnnals of Agricultural Sciences: Nur A. Zin, Noor A. BadaluddinAde regaNo ratings yet
- 2013 USSIRI - Soil Emission of Nitrous Oxide and Its MitigationDocument390 pages2013 USSIRI - Soil Emission of Nitrous Oxide and Its MitigationAnderson Dias SilveiraNo ratings yet
- Qualitative Test - Amino Acids and ProteinsDocument9 pagesQualitative Test - Amino Acids and Proteinstapans_8No ratings yet
- Group Presentation RubricDocument1 pageGroup Presentation RubricErika MatiasNo ratings yet
- Effects of Alternate Wetting and Drying Technique On Greenhouse Gas Emissions From Irrigated Rice Paddy in Central Luzon, PhilippinesDocument9 pagesEffects of Alternate Wetting and Drying Technique On Greenhouse Gas Emissions From Irrigated Rice Paddy in Central Luzon, PhilippinesErika MatiasNo ratings yet
- How To Solve Drug Dosage ProblemsDocument9 pagesHow To Solve Drug Dosage Problemspatra fadmaNo ratings yet
- Group Presentation RubricDocument1 pageGroup Presentation RubricErika MatiasNo ratings yet
- Washington State Cat Project Unit 3Document44 pagesWashington State Cat Project Unit 3CatalinAlbuNo ratings yet
- Moles, Units, and Conversion FactorsDocument19 pagesMoles, Units, and Conversion FactorsJermyn G EvangelistaNo ratings yet
- ORGANELLE Table PDFDocument2 pagesORGANELLE Table PDFDebjit BanerjeeNo ratings yet
- Prereg Drug Calculations Roy Sinclair Booklet 1Document66 pagesPrereg Drug Calculations Roy Sinclair Booklet 1Sudhakar KuppireddyNo ratings yet
- Structures of Amino AcidsDocument32 pagesStructures of Amino AcidsUsman GhaniNo ratings yet
- Mammalian digestive tract adaptationsDocument19 pagesMammalian digestive tract adaptationsErika MatiasNo ratings yet
- How Do Animals Breathe?: All Vertebrates Lungs DiaphragmDocument1 pageHow Do Animals Breathe?: All Vertebrates Lungs DiaphragmErika MatiasNo ratings yet
- The Nervous System 2Document84 pagesThe Nervous System 2Erika MatiasNo ratings yet
- The Integumentary Morphology of Modern BirdsDocument17 pagesThe Integumentary Morphology of Modern BirdsErika MatiasNo ratings yet
- Heart: Comparative Anatomy of Heart in Vertebrates Evolution of Heart in VertebratesDocument28 pagesHeart: Comparative Anatomy of Heart in Vertebrates Evolution of Heart in VertebratesErika MatiasNo ratings yet
- Aves SkinDocument7 pagesAves SkinErika MatiasNo ratings yet
- Comparative Anatomy of Heart in VertebratesDocument8 pagesComparative Anatomy of Heart in VertebratesErika MatiasNo ratings yet
- Comparative Anatomy of Heart in VertebratesDocument8 pagesComparative Anatomy of Heart in VertebratesErika MatiasNo ratings yet
- Circulatory System ComponentsDocument23 pagesCirculatory System ComponentsErika MatiasNo ratings yet
- Proline Protocol: Bates Et Al. (1973) : G Proline/ml X ML Toluene X G of SampleDocument2 pagesProline Protocol: Bates Et Al. (1973) : G Proline/ml X ML Toluene X G of SampleErika MatiasNo ratings yet
- Circulatory System ComponentsDocument23 pagesCirculatory System ComponentsErika MatiasNo ratings yet
- Mammalian digestive tract adaptationsDocument19 pagesMammalian digestive tract adaptationsErika MatiasNo ratings yet
- Horse Brain Human Brain-ExcerptDocument20 pagesHorse Brain Human Brain-ExcerptSruti Robee SNo ratings yet
- Animal AbuseDocument4 pagesAnimal Abuse3Q Junior Taman MidahNo ratings yet
- LH C1 Advanced - WBDocument96 pagesLH C1 Advanced - WBfelipegomez24041No ratings yet
- Komite Medik Rumah Sakit Islam GondanglegiDocument12 pagesKomite Medik Rumah Sakit Islam GondanglegiEry MufidaNo ratings yet
- Information On Leh Ladakh FestivalDocument7 pagesInformation On Leh Ladakh FestivalNisha AroraNo ratings yet
- Animal Farm - Knowledge OrganiserDocument2 pagesAnimal Farm - Knowledge OrganiserLabeenaNo ratings yet
- HematopoiesisDocument3 pagesHematopoiesisVino OrajayNo ratings yet
- Black Beauty Retold Story of Kindness and FreedomDocument22 pagesBlack Beauty Retold Story of Kindness and Freedomyupi_futbolNo ratings yet
- Dungeon #165Document100 pagesDungeon #165JackSchutte67% (3)
- Ultrasound Measurements Data Communication: Aloka Co., LTDDocument8 pagesUltrasound Measurements Data Communication: Aloka Co., LTDRama Tenis CopecNo ratings yet
- The Proud Deer's Lesson on HumilityDocument3 pagesThe Proud Deer's Lesson on Humilityyyee81No ratings yet
- Lowering Creatinine with Tagulinao Plant ExtractDocument70 pagesLowering Creatinine with Tagulinao Plant ExtractSean MarianoNo ratings yet
- Veterinary Practice Management SecretsDocument166 pagesVeterinary Practice Management SecretsMiguel Santos100% (6)
- Comparative Histologic Evaluation of Vertebrate OvDocument8 pagesComparative Histologic Evaluation of Vertebrate OvAndrea Zaragoza FernándezNo ratings yet
- Blood Questions Human Physiology PDFDocument5 pagesBlood Questions Human Physiology PDFARUSHI ARVIND100% (2)
- Serpentfolk: A Playable Race For 5 EditionDocument2 pagesSerpentfolk: A Playable Race For 5 EditionStefan CostaNo ratings yet
- Different Modes of Feeding and DigestionDocument4 pagesDifferent Modes of Feeding and Digestionczar0992No ratings yet
- Diuretic Effect and Mechanism of ActionDocument5 pagesDiuretic Effect and Mechanism of Actionwulan faddhylanNo ratings yet
- The Donkey Lady Fights La Llorona and Other Stories / La Señora Asna Se Enfrenta A La Llorona y Otros Cuentos by Xavier GarzaDocument186 pagesThe Donkey Lady Fights La Llorona and Other Stories / La Señora Asna Se Enfrenta A La Llorona y Otros Cuentos by Xavier GarzaArte Público PressNo ratings yet
- Different Types of Smokehouses and Smokehouse PlansDocument5 pagesDifferent Types of Smokehouses and Smokehouse PlansEloisa OleaNo ratings yet
- List of Dot Exempt CommotiesDocument31 pagesList of Dot Exempt CommotiesRa JacksonNo ratings yet
- Bab 2 Jurnal PriapismusDocument2 pagesBab 2 Jurnal PriapismusumarNo ratings yet
- Bud 2016 CidDocument351 pagesBud 2016 Cidgayan12001No ratings yet
- Titania of The Summer CourtDocument4 pagesTitania of The Summer CourtGeraldNo ratings yet
- Grammar Notes on Positive and Negative Statements, Articles, Quantifiers, Singular and Plural Nouns, and Possessive NounsDocument5 pagesGrammar Notes on Positive and Negative Statements, Articles, Quantifiers, Singular and Plural Nouns, and Possessive NounsNana ZainNo ratings yet
- Effects of Pumpkin and Bitter Leaf Extracts on Rat SpermDocument33 pagesEffects of Pumpkin and Bitter Leaf Extracts on Rat SpermThompson E IghaloNo ratings yet
- Basic 5 Second Term ExamDocument11 pagesBasic 5 Second Term ExamDaniel Bamgbose100% (1)
- Lecture 5 Nutrient Requirement of Laying HensDocument36 pagesLecture 5 Nutrient Requirement of Laying Hensumi amirah100% (1)