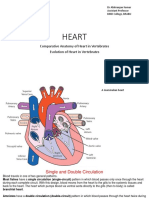
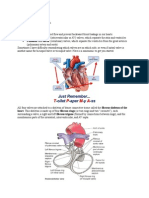
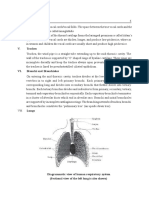
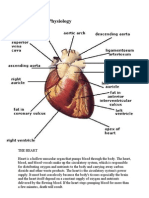
You might also like
- Evolution of Heart in VertebratesDocument4 pagesEvolution of Heart in Vertebratesनितिन कुमारNo ratings yet
- Heart: Comparative Anatomy of Heart in Vertebrates Evolution of Heart in VertebratesDocument28 pagesHeart: Comparative Anatomy of Heart in Vertebrates Evolution of Heart in VertebratesErika MatiasNo ratings yet
- The Structure and Function of the Heart Across the Animal KingdomDocument9 pagesThe Structure and Function of the Heart Across the Animal KingdomDanagalla Venkat100% (2)
- Circulatory System in Finfish and ShellfishDocument27 pagesCirculatory System in Finfish and ShellfishDr. Abir IshtiaqNo ratings yet
- In FishDocument1 pageIn Fishscribd388No ratings yet
- Cardiovascular System - Heart: AssignmentDocument14 pagesCardiovascular System - Heart: AssignmentKriss MisraNo ratings yet
- Transport in Invertebrates and VertebratesDocument13 pagesTransport in Invertebrates and VertebratesJoan GalarceNo ratings yet
- U6C6 - Carty Soriano - Heart ValvesDocument6 pagesU6C6 - Carty Soriano - Heart ValvesXio LinaresNo ratings yet
- Anatomy and Physiology of HeartDocument69 pagesAnatomy and Physiology of HeartRitu Arun Kaushik100% (1)
- The Fully Divided HeartDocument1 pageThe Fully Divided HeartSulochana ChanNo ratings yet
- The Cardiovascular System The HeartDocument9 pagesThe Cardiovascular System The HeartMarjorie AbelgasNo ratings yet
- Subject Enrichment Activity For Term 2Document5 pagesSubject Enrichment Activity For Term 2AADITRYA JAINNo ratings yet
- Circulatory System in FinfishDocument18 pagesCirculatory System in FinfishSalma SabirNo ratings yet
- Paul Cardiac Lecture 2015 OutlineDocument6 pagesPaul Cardiac Lecture 2015 OutlineMichalis KantartzisNo ratings yet
- Anatomy of The HeartDocument5 pagesAnatomy of The HeartxXjhmXxNo ratings yet
- Gross Anatomy II Lecture on Heart and Vascular SystemDocument13 pagesGross Anatomy II Lecture on Heart and Vascular SystemSantosh Bhandari100% (2)
- Circulatory SystemDocument111 pagesCirculatory SystemYounas Bhatti100% (1)
- CardiovascularDocument92 pagesCardiovascularRolinette DaneNo ratings yet
- ANP Heart Anatomy & PhysiologyDocument3 pagesANP Heart Anatomy & PhysiologySheana Tmpl100% (1)
- V. Trachea: Zoology-IIDocument22 pagesV. Trachea: Zoology-IIVenkatesh JoguNo ratings yet
- Anatomy and Physiology Lecture NotesDocument6 pagesAnatomy and Physiology Lecture NotesBlackcat KememeyNo ratings yet
- Circulation Vocabulary HandoutDocument2 pagesCirculation Vocabulary HandoutGPearce204No ratings yet
- Campbell Chapter 42 NotesDocument17 pagesCampbell Chapter 42 NotesRyan LiuNo ratings yet
- Jantung (The Heart)Document24 pagesJantung (The Heart)JoePutraNo ratings yet
- Heart Anatomy ExplainedDocument15 pagesHeart Anatomy ExplainedAritra SahaNo ratings yet
- MS Wrep IIDocument51 pagesMS Wrep IIiana-almocera-6970No ratings yet
- The Heart: Anatomy, Chambers, Circulation & ValvesDocument14 pagesThe Heart: Anatomy, Chambers, Circulation & ValvesGary Bowen100% (1)
- The Circulatory System: Transport for CellsDocument4 pagesThe Circulatory System: Transport for CellsPuguh HadiNo ratings yet
- Anatomy of the Heart - Layers, Chambers, ValvesDocument28 pagesAnatomy of the Heart - Layers, Chambers, ValvesHerman DjawaNo ratings yet
- Anatomy and Histology of The Cardiovascular SystemDocument16 pagesAnatomy and Histology of The Cardiovascular SystemTasmiah HossainNo ratings yet
- DFF FFF AaaaaDocument5 pagesDFF FFF AaaaaffffffffNo ratings yet
- ССС 2019Document83 pagesССС 2019bekafop813No ratings yet
- Anatomy and Physiology of the Heart ExplainedDocument6 pagesAnatomy and Physiology of the Heart ExplainedChamCham AquinoNo ratings yet
- The Circulatory System in Man: A Concise Guide to the Heart and Blood FlowDocument54 pagesThe Circulatory System in Man: A Concise Guide to the Heart and Blood FlowStudent ResearchNo ratings yet
- HUMAN ANATOMY AND PHYSIOLOGY LABORATORY Cardiovascular SystemDocument12 pagesHUMAN ANATOMY AND PHYSIOLOGY LABORATORY Cardiovascular SystemZaira MangalimanNo ratings yet
- CH 6 - The Circulatory System PDFDocument11 pagesCH 6 - The Circulatory System PDFAnonymous e6LINHNo ratings yet
- Functions of Cardiovascular System:: The Gross AnatomyDocument6 pagesFunctions of Cardiovascular System:: The Gross AnatomyShreyasi PatankarNo ratings yet
- Animals Part 1Document129 pagesAnimals Part 1黄心怡No ratings yet
- Chapter 13 - HeartDocument5 pagesChapter 13 - Heartbussniesm05No ratings yet
- The Circulatory SystemDocument8 pagesThe Circulatory SystembysshelarbyNo ratings yet
- Functions of the HeartDocument15 pagesFunctions of the HeartAngelica UrbiNo ratings yet
- The Circulatory System: Agriscience 332 Animal Science #8646-A TEKS: (C) (2) (A) and (C) (2) (B)Document111 pagesThe Circulatory System: Agriscience 332 Animal Science #8646-A TEKS: (C) (2) (A) and (C) (2) (B)brdaisyNo ratings yet
- Notes Grade: 9 Subject: Biology Chapter No: 9 Chapter Name: Transport in Animals ObjectivesDocument36 pagesNotes Grade: 9 Subject: Biology Chapter No: 9 Chapter Name: Transport in Animals Objectivespraneeth satishNo ratings yet
- Anatomy and Physiology of The HeartDocument5 pagesAnatomy and Physiology of The HeartMuri CajucomNo ratings yet
- 7.8.heart and VesselsDocument11 pages7.8.heart and Vesselssomebody_maNo ratings yet
- Cardiovascular System DR Aarti Abhishek Shah NmimsDocument33 pagesCardiovascular System DR Aarti Abhishek Shah NmimsnoshNo ratings yet
- Angiology Lecture 2Document5 pagesAngiology Lecture 2আহানাফ তাহমিদ শব্দNo ratings yet
- PHYSIOLOGY OF THE CARDIOVASCULAR SYSTEMDocument92 pagesPHYSIOLOGY OF THE CARDIOVASCULAR SYSTEMSanti Purnama SariNo ratings yet
- Internal Structure of The Heart - Contemporary Health IssuesDocument1 pageInternal Structure of The Heart - Contemporary Health IssuesMax RamirezNo ratings yet
- Circulatory 20Pathways-STUDENTDocument53 pagesCirculatory 20Pathways-STUDENTMark M. AlipioNo ratings yet
- Internal Structure of HeartDocument1 pageInternal Structure of Heartmbudhathoki3000No ratings yet
- Heart: StructureDocument11 pagesHeart: StructureKert trocioNo ratings yet
- Cardiovascular PBLsDocument8 pagesCardiovascular PBLsAya SobhiNo ratings yet
- Transport in Man NotesDocument9 pagesTransport in Man NotesYeo JhNo ratings yet
- Cardiovascular and Respiratory SystemDocument3 pagesCardiovascular and Respiratory SystemGodsend OparaNo ratings yet
- Anatomy Physiology HeartDocument9 pagesAnatomy Physiology HeartAlvheen JoaquinNo ratings yet
- Project BioDocument18 pagesProject Bioyanshu falduNo ratings yet
- Circulatory System: A Tutorial Study GuideFrom EverandCirculatory System: A Tutorial Study GuideRating: 5 out of 5 stars5/5 (3)
- 7.3 Mechanical DigestionDocument5 pages7.3 Mechanical DigestionelizabethNo ratings yet
- Basic Computer Quiz - MCQ on Components, Generations & HistoryDocument7 pagesBasic Computer Quiz - MCQ on Components, Generations & Historyprem sagar100% (2)
- CopyofCopyofMaldeepSingh Jawanda ResumeDocument2 pagesCopyofCopyofMaldeepSingh Jawanda Resumebob nioNo ratings yet
- Viennot - 1979 - Spontaneous Reasoning in Elementary DynamicsDocument18 pagesViennot - 1979 - Spontaneous Reasoning in Elementary Dynamicsjumonteiro2000No ratings yet
- 41-How To Calculate Air Temp in Unconditioned SpacesDocument3 pages41-How To Calculate Air Temp in Unconditioned Spacesalmig200No ratings yet
- WhatsoldDocument141 pagesWhatsoldLuciana KarajalloNo ratings yet
- Nad C541iDocument37 pagesNad C541iapi-3837207No ratings yet
- Hele 4 PPT - 2nd QT - Week 1Document47 pagesHele 4 PPT - 2nd QT - Week 1jerico juarezNo ratings yet
- Creatinine JaffeDocument2 pagesCreatinine JaffeOsinachi WilsonNo ratings yet
- Few Words About Digital Protection RelayDocument5 pagesFew Words About Digital Protection RelayVasudev AgrawalNo ratings yet
- (Nano and Energy) Gavin Buxton - Alternative Energy Technologies - An Introduction With Computer Simulations-CRC Press (2014) PDFDocument302 pages(Nano and Energy) Gavin Buxton - Alternative Energy Technologies - An Introduction With Computer Simulations-CRC Press (2014) PDFmarcosNo ratings yet
- HE HOUSEKEEPING GR11 Q1 MODULE-6-for-teacherDocument25 pagesHE HOUSEKEEPING GR11 Q1 MODULE-6-for-teacherMikaela YtacNo ratings yet
- Consumer Behaviour of Titan WatchesDocument57 pagesConsumer Behaviour of Titan Watchesmanu100% (1)
- Ultra Structure of Plant Cell 1Document18 pagesUltra Structure of Plant Cell 1Kumaran JothiramNo ratings yet
- Panasonic SA-HT878Document82 pagesPanasonic SA-HT878immortalwombatNo ratings yet
- GI6.020 Personal Flotation Devices (PFDS) 10-31-2009Document5 pagesGI6.020 Personal Flotation Devices (PFDS) 10-31-2009GooglegulNo ratings yet
- CENELEC RA STANDARDS CATALOGUEDocument17 pagesCENELEC RA STANDARDS CATALOGUEHamed AhmadnejadNo ratings yet
- TaxonomyDocument56 pagesTaxonomyKrezia Mae SolomonNo ratings yet
- Steady State Modeling and Simulation of The Oleflex Process For Isobutane Dehydrogenation Considering Reaction NetworkDocument9 pagesSteady State Modeling and Simulation of The Oleflex Process For Isobutane Dehydrogenation Considering Reaction NetworkZangNo ratings yet
- 2024 Yoga Vidya Training FormDocument8 pages2024 Yoga Vidya Training FormJohnNo ratings yet
- MetalCut User ManualDocument71 pagesMetalCut User Manualnammaris0% (1)
- Letter of Invitation ESVC 3000 2024 1Document2 pagesLetter of Invitation ESVC 3000 2024 1pradeep kumarNo ratings yet
- 00001Document20 pages00001Maggie ZhuNo ratings yet
- DMG48480F021 01WN DataSheetDocument16 pagesDMG48480F021 01WN DataSheeteminkiranNo ratings yet
- The History of Coins and Banknotes in Mexico: September 2012Document35 pagesThe History of Coins and Banknotes in Mexico: September 2012Mladen VidovicNo ratings yet
- BLADED - Theory Manual PDFDocument134 pagesBLADED - Theory Manual PDFdavidlokito100% (2)
- Year 7 Bugs Lesson 2Document1 pageYear 7 Bugs Lesson 2api-293503824No ratings yet
- Method Statement For Planter WaterproofingDocument8 pagesMethod Statement For Planter WaterproofingMonali Varpe0% (1)
- Audiology DissertationDocument4 pagesAudiology DissertationPaperWritingHelpOnlineUK100% (1)