You might also like
- Pedigree Worksheet Answer KeyDocument1 pagePedigree Worksheet Answer Keyapi-477608730100% (8)
- Advanced Assessment Interpreting Findings and Formulating Differential Diagnoses 3rd Edition Goolsby Test BankDocument9 pagesAdvanced Assessment Interpreting Findings and Formulating Differential Diagnoses 3rd Edition Goolsby Test BankAlbertBradyypies100% (16)
- What is Genetics in 40 CharactersDocument3 pagesWhat is Genetics in 40 Characterskevin maravilla100% (3)
- Rubric Genetic Disorder ProjectDocument4 pagesRubric Genetic Disorder Projectajl5010No ratings yet
- General Principles of Hereditary DiseasesDocument17 pagesGeneral Principles of Hereditary DiseasesrutwickNo ratings yet
- Pharmacology Course for Nursing StudentsDocument15 pagesPharmacology Course for Nursing Studentssingireddi1revathi100% (2)
- Referat Thalasemia MajorDocument21 pagesReferat Thalasemia Majoreryprayudi13No ratings yet
- Standards and EhealthDocument20 pagesStandards and EhealthITU-T Technology Watch50% (2)
- Gen SchizopDocument11 pagesGen SchizopDewiakyuNo ratings yet
- Gordon Et Al - 2019Document15 pagesGordon Et Al - 2019Chrisjesse NwachukwuNo ratings yet
- Mutations in Potassium Channel Kir2.6 Cause Susceptibility To Thyrotoxic Hypokalemic Periodic ParalysisDocument11 pagesMutations in Potassium Channel Kir2.6 Cause Susceptibility To Thyrotoxic Hypokalemic Periodic ParalysisVenyLieNo ratings yet
- Gim 201618Document8 pagesGim 201618Ioana PoterasNo ratings yet
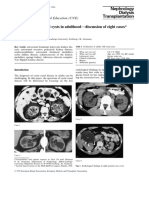
- Polycystic Kidney - Eight CasesDocument11 pagesPolycystic Kidney - Eight CasesKAINAT ANJUMNo ratings yet
- Genetics NotesDocument5 pagesGenetics NotesmidegeNo ratings yet
- Recurrent de Novo Point Mutations in Lamin A Cause Hutchinson-Gilford Progeria SyndromeDocument6 pagesRecurrent de Novo Point Mutations in Lamin A Cause Hutchinson-Gilford Progeria SyndromeGho VinsenNo ratings yet
- ParkinsonDocument9 pagesParkinsonAnonymous lwYEylQhNo ratings yet
- Il 7Document6 pagesIl 7Shukr Wesman BlbasNo ratings yet
- JHG 201178Document3 pagesJHG 201178DAVID TERRONESNo ratings yet
- Digenic Parkinsonism: Investigation of The Synergistic Effects of PRKN and LRRK2Document5 pagesDigenic Parkinsonism: Investigation of The Synergistic Effects of PRKN and LRRK2SakinaNo ratings yet
- AP1S2 Is Mutated in Xlinked DandyWalker Malformation With Intellectual Disability Basal Ganglia Disease and Seizures Pettigrew Syndrome - 2014Document6 pagesAP1S2 Is Mutated in Xlinked DandyWalker Malformation With Intellectual Disability Basal Ganglia Disease and Seizures Pettigrew Syndrome - 2014Jose Rafael Villafan BernalNo ratings yet
- A Dup (17) (p11.2p11.2) Detected by Fluorescence in Situ Hybridization in A Boy With Alport SyndromeDocument4 pagesA Dup (17) (p11.2p11.2) Detected by Fluorescence in Situ Hybridization in A Boy With Alport SyndromeRajan PandaNo ratings yet
- Srinivasan 2016Document16 pagesSrinivasan 2016Zeljko TomljanovicNo ratings yet
- Natural Selection at The RASGEF1C (GGC) Repeat in Human and Divergent Genotypes in Late Onset Neurocognitive DisorderDocument8 pagesNatural Selection at The RASGEF1C (GGC) Repeat in Human and Divergent Genotypes in Late Onset Neurocognitive DisorderAnahí TessaNo ratings yet
- Science 1997Document3 pagesScience 1997demmie1989No ratings yet
- Duplication at Xq13.3-q21.1 With Syndromic Intellectual Disability, A Probable Role For The ATRX GeneDocument6 pagesDuplication at Xq13.3-q21.1 With Syndromic Intellectual Disability, A Probable Role For The ATRX GeneEyyup UctepeNo ratings yet
- TMP 29 CDDocument15 pagesTMP 29 CDFrontiersNo ratings yet
- Aicardi-Goutie'res Syndrome: Neuroradiologic Findings and Follow-UpDocument6 pagesAicardi-Goutie'res Syndrome: Neuroradiologic Findings and Follow-UpclaypotgoldNo ratings yet
- Biopolym - Cell 2018 34 5 361 enDocument6 pagesBiopolym - Cell 2018 34 5 361 enАнна ШаповаловаNo ratings yet
- Biopolym - Cell 2018 34 5 361 enDocument6 pagesBiopolym - Cell 2018 34 5 361 enАнна ШаповаловаNo ratings yet
- Co Occurrence of Sporadic Parkinsonism ADocument4 pagesCo Occurrence of Sporadic Parkinsonism ARaja Som RajNo ratings yet
- Tuberous Sclerosis Complex: Advances in Diagnosis, Genetics, and ManagementDocument14 pagesTuberous Sclerosis Complex: Advances in Diagnosis, Genetics, and ManagementAri KurniawanNo ratings yet
- Molecular Genetic Analysis of Steroid Resistant NephroticDocument8 pagesMolecular Genetic Analysis of Steroid Resistant NephroticBeatri AyuzaNo ratings yet
- LupusDocument7 pagesLupusZyanCeronNo ratings yet
- Silver Russell 2010Document8 pagesSilver Russell 2010Jorge SHNo ratings yet
- Nob 30093Document7 pagesNob 30093Bilel Le ChevalierNo ratings yet
- PKAN NEJMDocument8 pagesPKAN NEJMmartin.ruda93No ratings yet
- Genotype-Phenotype Correlation, and Phenotypic HeterogeneityDocument9 pagesGenotype-Phenotype Correlation, and Phenotypic Heterogeneitykorokoro12No ratings yet
- TMP 756 DDocument24 pagesTMP 756 DnithiananthiNo ratings yet
- Sindrome de GitelmanDocument7 pagesSindrome de GitelmanHenry Perugachi MendezNo ratings yet
- 1737 FullDocument5 pages1737 Fullnurul auliaNo ratings yet
- Phacomatosis spilosebacea: A new name for a distinctive binary genodermatosisDocument10 pagesPhacomatosis spilosebacea: A new name for a distinctive binary genodermatosisPedroNo ratings yet
- Synaptic Ubiquitin Ligases: 706 Cell 147, October 28, 2011 ©2011 Elsevier Inc. DOI 10.1016/j.cell.2011.10.015Document2 pagesSynaptic Ubiquitin Ligases: 706 Cell 147, October 28, 2011 ©2011 Elsevier Inc. DOI 10.1016/j.cell.2011.10.015LivsNo ratings yet
- Sindrome NefroticoDocument8 pagesSindrome NefroticoRudy Espejo del ArcaNo ratings yet
- DNA Methylome R-Loop and Clinical Exome ProfilingDocument13 pagesDNA Methylome R-Loop and Clinical Exome ProfilingShopnil SarkarNo ratings yet
- Sinnecker 1996Document8 pagesSinnecker 1996Daniela Pereira LaureanoNo ratings yet
- The Landscape of Epilepsy-Related GATOR1 Variants Baldassari 2019Document11 pagesThe Landscape of Epilepsy-Related GATOR1 Variants Baldassari 2019cicasmileNo ratings yet
- EEG en Creuzfeld JacobDocument17 pagesEEG en Creuzfeld JacobKike jovelNo ratings yet
- ElderlyDocument1 pageElderlyapi-26340035No ratings yet
- 2020 Clinical Association of White MatterDocument9 pages2020 Clinical Association of White MatterMarco MerazNo ratings yet
- AAV9 - s41591 019 0674 1Document39 pagesAAV9 - s41591 019 0674 1Felipe CourtNo ratings yet
- Grant's Atlas of Anatomy Grant's Dissector: JAMA The Journal of The American Medical Association January 2008Document6 pagesGrant's Atlas of Anatomy Grant's Dissector: JAMA The Journal of The American Medical Association January 2008valmondsikiNo ratings yet
- 2008 - MarshallDocument12 pages2008 - MarshallCarlosNo ratings yet
- Gene expression differences between thyroid tumorsDocument11 pagesGene expression differences between thyroid tumorsArtha MahulaeNo ratings yet
- Structural Brain and Spinal Cord Damage in Symptomatic and Pre-Symptomatic VAPB-related ALSpdfDocument6 pagesStructural Brain and Spinal Cord Damage in Symptomatic and Pre-Symptomatic VAPB-related ALSpdfDanilo SilvaNo ratings yet
- IL-7R α polymorphisms in 60 Iranian multiple sclerosis patientsDocument6 pagesIL-7R α polymorphisms in 60 Iranian multiple sclerosis patientsShukr Wesman BlbasNo ratings yet
- Evidence For A Common Spinocerebellar Ataxia Type 7: (SCA7) Founder Mutation in ScandinaviaDocument5 pagesEvidence For A Common Spinocerebellar Ataxia Type 7: (SCA7) Founder Mutation in ScandinaviaJuliana SiruffoNo ratings yet
- Paper TEADocument10 pagesPaper TEAmetteoroNo ratings yet
- Aceves-Ramirez2022Document8 pagesAceves-Ramirez2022Diana MartinezNo ratings yet
- 27 PDFDocument9 pages27 PDFMohamad FakihNo ratings yet
- Association of MASP2 Levels and MASP2 Gene Polymorphisms With Systemic Lupus ErythematosusDocument12 pagesAssociation of MASP2 Levels and MASP2 Gene Polymorphisms With Systemic Lupus ErythematosusShjajaNo ratings yet
- 1 s2.0 S038776040900299X MainDocument6 pages1 s2.0 S038776040900299X MainLeila Schuindt MonneratNo ratings yet
- 1 s2.0 S0002929707624161 Main PDFDocument8 pages1 s2.0 S0002929707624161 Main PDFaris ekoNo ratings yet
- Jurnal Rett SyndromeDocument7 pagesJurnal Rett SyndromeFitri IsnainiNo ratings yet
- BrainCellsDifferencesScience2015 PDFDocument6 pagesBrainCellsDifferencesScience2015 PDFJose Carlos Alarcon MendezNo ratings yet
- Sazci2012. Gen ZNF804ADocument5 pagesSazci2012. Gen ZNF804AFahrunnisa NurdinNo ratings yet
- Artículo ViroDocument13 pagesArtículo ViroNatalia BogotáNo ratings yet
- PpokDocument7 pagesPpokSkoy KuyNo ratings yet
- Cavalli 2009Document6 pagesCavalli 2009Ifany oschaliaNo ratings yet
- Genmed HtsDocument17 pagesGenmed HtsIfany oschaliaNo ratings yet
- Genetic DiagnosaDocument6 pagesGenetic DiagnosaIfany oschaliaNo ratings yet
- Wiggin Ton 2005Document7 pagesWiggin Ton 2005Ifany oschaliaNo ratings yet
- Crow 1999Document5 pagesCrow 1999Ifany oschaliaNo ratings yet
- Common Paediatric Genetic Disorders - CUHK - Paed - 2018Document21 pagesCommon Paediatric Genetic Disorders - CUHK - Paed - 2018B AuNo ratings yet
- Toronto Sick KidsDocument13 pagesToronto Sick KidsAnonymous AuJncFVWvNo ratings yet
- Biochemistry of Down SyndromDocument3 pagesBiochemistry of Down SyndromJosephine mbahNo ratings yet
- M3 WHILE TASK For Lab Exercise 10Document3 pagesM3 WHILE TASK For Lab Exercise 10Michelle LutapNo ratings yet
- Medical Genetics: Assoc Prof Rusu Cristina, MD PHDDocument23 pagesMedical Genetics: Assoc Prof Rusu Cristina, MD PHDRox CatalinaNo ratings yet
- Final RET Bulletin - 2020-2021Document58 pagesFinal RET Bulletin - 2020-2021ADESH MISHRANo ratings yet
- Introduction To Medical Genetics Lecture NotesDocument41 pagesIntroduction To Medical Genetics Lecture NotesFarhan KhosaNo ratings yet
- Ukg Asahan - ExcelDocument124 pagesUkg Asahan - ExcelSutantoNo ratings yet
- Genetic Counselling GuideDocument14 pagesGenetic Counselling Guidetuppadb50% (2)
- Committee On Veterans' Affairs House of Representatives: HearingDocument113 pagesCommittee On Veterans' Affairs House of Representatives: HearingScribd Government DocsNo ratings yet
- AchondroplasiaDocument3 pagesAchondroplasiaramziaNo ratings yet
- Lupin Vs Horizon Ravicti '559 Patent IPR Final Written DecisionDocument33 pagesLupin Vs Horizon Ravicti '559 Patent IPR Final Written DecisionTradeHawkNo ratings yet
- Muscular DystrophyDocument14 pagesMuscular Dystrophyapi-439738433No ratings yet
- My PG Notes® - Disorders With Café-Au-Lait SpotsDocument1 pageMy PG Notes® - Disorders With Café-Au-Lait SpotsSubhajitPaulNo ratings yet
- Fellowship in Genetic Diagnostics for Government CliniciansDocument8 pagesFellowship in Genetic Diagnostics for Government Cliniciansm nNo ratings yet
- Introduction To HemoglobinopathiesDocument12 pagesIntroduction To HemoglobinopathiesMichael MazvidzaNo ratings yet
- SOP For WEBDocument18 pagesSOP For WEBhasanNo ratings yet
- What Is AdermatoglyphiaDocument2 pagesWhat Is AdermatoglyphiaAdriel C LibanganNo ratings yet
- Clinical Key Master - Content - List - 14 Jun 2019Document49 pagesClinical Key Master - Content - List - 14 Jun 2019Axevim GNo ratings yet
- CCMG General Knowledge Study Guide 2017Document7 pagesCCMG General Knowledge Study Guide 2017Md Hasan ImamNo ratings yet
- Genetic Disorder Brochure ProjectDocument1 pageGenetic Disorder Brochure ProjectErika ContrerasNo ratings yet
- Abruzzo-Erickson SyndromeDocument3 pagesAbruzzo-Erickson SyndromeMario Luis LuquezNo ratings yet