You might also like
- Clinical Ophthalmic Oncology: RetinoblastomaFrom EverandClinical Ophthalmic Oncology: RetinoblastomaJesse L. BerryNo ratings yet
- Prenatal Detection of Congenital Anomalies and Related Factors in ArgentinaDocument10 pagesPrenatal Detection of Congenital Anomalies and Related Factors in Argentinajorge davidNo ratings yet
- Clinical Nephrogeriatrics: An Evidence-Based GuideFrom EverandClinical Nephrogeriatrics: An Evidence-Based GuideCarlos Guido MussoNo ratings yet
- Alteraciones Cromosomicas Molecular GeneticsDocument14 pagesAlteraciones Cromosomicas Molecular GeneticsChristian AguirreNo ratings yet
- Interferon Regulatory Factor 6 Gene Variants and The Risk of Isolated Cleft Lip or PalateDocument12 pagesInterferon Regulatory Factor 6 Gene Variants and The Risk of Isolated Cleft Lip or PalateDwitiara SeptianiNo ratings yet
- NIH Public Access: Author ManuscriptDocument12 pagesNIH Public Access: Author ManuscriptJosé EstevesNo ratings yet
- Thornton2018 ANGI Methods ContempClinicalTrials74 p61Document24 pagesThornton2018 ANGI Methods ContempClinicalTrials74 p61treby NaterNo ratings yet
- Vazquez-Mojena-13 Epidemio HD CubaDocument4 pagesVazquez-Mojena-13 Epidemio HD CubaMARIA MONTSERRAT SOMOZA MONCADANo ratings yet
- Exome Sequencing in Multiplex Autism Families Suggests A Major Role For Heterozygous Truncating MutationsDocument7 pagesExome Sequencing in Multiplex Autism Families Suggests A Major Role For Heterozygous Truncating MutationsTrúc LêNo ratings yet
- 2013 Clinical WES Nejmoa1306555Document10 pages2013 Clinical WES Nejmoa1306555MCuk2606No ratings yet
- Retinoblastoma Update Pediatric Eye TumorDocument10 pagesRetinoblastoma Update Pediatric Eye TumorTatiana RecinosNo ratings yet
- Original ArticleDocument7 pagesOriginal ArticleDain 2IronfootNo ratings yet
- Nasolaryngoscopy in A Family Medicine Clinic: Indications, Findings, and EconomicsDocument7 pagesNasolaryngoscopy in A Family Medicine Clinic: Indications, Findings, and Economicsriski novitaNo ratings yet
- Pestisida Dan AnensefaliDocument9 pagesPestisida Dan AnensefaliMaman AbdurahmanNo ratings yet
- Neurocisticercosis en Una Población Pediátrica en Lima: Análisis Epidemiológico y ClínicoDocument4 pagesNeurocisticercosis en Una Población Pediátrica en Lima: Análisis Epidemiológico y ClínicoReyna del PilarNo ratings yet
- Murgueitio Et Al., 2023 Importance of Including Individuals of Latin American Ancestry in Genetic Studies of Feeding and Eating Disorders.Document22 pagesMurgueitio Et Al., 2023 Importance of Including Individuals of Latin American Ancestry in Genetic Studies of Feeding and Eating Disorders.Emilio J. CompteNo ratings yet
- Sickle Cell DiseaseDocument10 pagesSickle Cell Diseasekarthikavvari2002No ratings yet
- Riley 2009Document9 pagesRiley 2009Fahrunnisa NurdinNo ratings yet
- Epilepsy-Asia .Review Lancet Neurol 2007Document11 pagesEpilepsy-Asia .Review Lancet Neurol 2007fikaNo ratings yet
- Faisal Usman SynopsisDocument6 pagesFaisal Usman SynopsisRahid KhanNo ratings yet
- tmp6A3C TMPDocument28 pagestmp6A3C TMPFrontiersNo ratings yet
- CMT WEGS Nejmoa0908094Document11 pagesCMT WEGS Nejmoa0908094MCuk2606No ratings yet
- The Mouse As A Model For Understanding Chronic Diseases of Aging: The Histopathologic Basis of Aging in Inbred MiceDocument9 pagesThe Mouse As A Model For Understanding Chronic Diseases of Aging: The Histopathologic Basis of Aging in Inbred MiceashorammarNo ratings yet
- Barbados Study1994Document9 pagesBarbados Study1994JULIO COLAN MORANo ratings yet
- Prader WilliDocument15 pagesPrader WilliGVHHNo ratings yet
- Medina Et Al-2005-EpilepsiaDocument8 pagesMedina Et Al-2005-EpilepsiaGriselNo ratings yet
- Incidence and Prevalence of Acute Lymphoblastic Leukemia (All) in Kashmiri Populace (North India)Document6 pagesIncidence and Prevalence of Acute Lymphoblastic Leukemia (All) in Kashmiri Populace (North India)IJAR JOURNALNo ratings yet
- 2017 Harripaul ColdSpringHarbPerspectMedDocument14 pages2017 Harripaul ColdSpringHarbPerspectMedEAPNo ratings yet
- Logroscino2019 SlaDocument11 pagesLogroscino2019 Slab.modNo ratings yet
- Artigo YFDocument15 pagesArtigo YFJanaina JohnssonNo ratings yet
- Dlova Et Al-2018-British Journal of DermatologyDocument2 pagesDlova Et Al-2018-British Journal of DermatologyAndrea CordaroNo ratings yet
- Zheng Hong 郑 红 Department of Medical Genetics & Cell BiologyDocument63 pagesZheng Hong 郑 红 Department of Medical Genetics & Cell BiologyinakiNo ratings yet
- New England Journal Medicine: The ofDocument10 pagesNew England Journal Medicine: The ofSuwandi ChangNo ratings yet
- Autismo EgiptoDocument8 pagesAutismo EgiptoOvsd Vfsg SDNo ratings yet
- Artigo C Neoformans PDFDocument29 pagesArtigo C Neoformans PDFJorge Luis CarmoNo ratings yet
- Genome Pathways For Cell-09-643644Document12 pagesGenome Pathways For Cell-09-643644AlmedinaNo ratings yet
- 5-ensuring-equity-diversity-and-inclusiveness-in-genetic-analysis-will-empower-the-future-ofDocument4 pages5-ensuring-equity-diversity-and-inclusiveness-in-genetic-analysis-will-empower-the-future-ofabdeali hazariNo ratings yet
- Clinical and Genetic Characteristics of Xeroderma Pigmentosum in NepalDocument18 pagesClinical and Genetic Characteristics of Xeroderma Pigmentosum in NepalKandy RinconNo ratings yet
- Guidelines For Case Classification For The National Birth Defects Prevention StudyDocument9 pagesGuidelines For Case Classification For The National Birth Defects Prevention Studyjorge davidNo ratings yet
- Movement Disorders - 2024 - Gan or - Genetics of Parkinson S Disease in Underrepresented Populations New Studies Pave TheDocument2 pagesMovement Disorders - 2024 - Gan or - Genetics of Parkinson S Disease in Underrepresented Populations New Studies Pave TheHenrique OliveiraNo ratings yet
- Project PaperDocument12 pagesProject Paperop gamerNo ratings yet
- A Systematic Review of Secretory Carcinoma of The Salivary Gland: Where Are We?Document10 pagesA Systematic Review of Secretory Carcinoma of The Salivary Gland: Where Are We?Uriel EnriquezNo ratings yet
- Ankylosing SpondylitesDocument16 pagesAnkylosing SpondylitesJyotirmayeeNo ratings yet
- Genetics of Autism Project Launches with 6,000 Sample Data SetDocument2 pagesGenetics of Autism Project Launches with 6,000 Sample Data Setemma199No ratings yet
- Ophthalmogenetic and Epidemiological Studies of Egyptian Children With Mental RetardationDocument8 pagesOphthalmogenetic and Epidemiological Studies of Egyptian Children With Mental Retardationray m deraniaNo ratings yet
- UKPMC Funders Group: Author ManuscriptDocument13 pagesUKPMC Funders Group: Author ManuscriptSharon Alva DominguezNo ratings yet
- Ajmg A 61300Document9 pagesAjmg A 61300Nigel GarayNo ratings yet
- 2011 - CooperDocument11 pages2011 - CooperCarlNo ratings yet
- Artificial Intelligence in Ovarian Cancer HistopatDocument14 pagesArtificial Intelligence in Ovarian Cancer Histopatbrolyssj088No ratings yet
- Clinico-Etiological Spectrum of Pancytopenia in Hospitalized ChildrenDocument4 pagesClinico-Etiological Spectrum of Pancytopenia in Hospitalized ChildrenNurul Huda KowitaNo ratings yet
- 2010 - Epidemioloy of Ulcerative Keratitis in Northern CaliforniaDocument7 pages2010 - Epidemioloy of Ulcerative Keratitis in Northern CaliforniaPrasetya AnugrahNo ratings yet
- File DementiaDocument12 pagesFile DementiaLucienneNo ratings yet
- Aicardi-Goutie'res Syndrome: Neuroradiologic Findings and Follow-UpDocument6 pagesAicardi-Goutie'res Syndrome: Neuroradiologic Findings and Follow-UpclaypotgoldNo ratings yet
- A Genetics-First Approach To Dissecting The Heterogeneity of Autism Phenotypic Comparison of Autism Risk Copy Number VariantsDocument27 pagesA Genetics-First Approach To Dissecting The Heterogeneity of Autism Phenotypic Comparison of Autism Risk Copy Number VariantsTiago ChavesNo ratings yet
- A Descriptive Study of NPHS1 and NPHS2 Mutations in ChildrenDocument14 pagesA Descriptive Study of NPHS1 and NPHS2 Mutations in ChildrenCalvin Tanuwijaya Stick BolaNo ratings yet
- Factores en Niños GuatemalaDocument9 pagesFactores en Niños GuatemalaFernando OrozcoNo ratings yet
- Rev Chil Pediatr 2007Document10 pagesRev Chil Pediatr 2007Skarleet PaolaaNo ratings yet
- Cause-Specific Child Mortality In A Mountainous Pakistani Community By Verbal AutopsyDocument8 pagesCause-Specific Child Mortality In A Mountainous Pakistani Community By Verbal AutopsyDr. Muhammad Bilal SiddiquiNo ratings yet
- Psychiatry Clin Neurosci - 2017 - Waye - Genetics and Epigenetics of Autism A ReviewDocument17 pagesPsychiatry Clin Neurosci - 2017 - Waye - Genetics and Epigenetics of Autism A ReviewKaline DantasNo ratings yet
- Optic Nerve Glioma: Case Series With Review of Clinical, Radiologic, Molecular, and Histopathologic CharacteristicsDocument5 pagesOptic Nerve Glioma: Case Series With Review of Clinical, Radiologic, Molecular, and Histopathologic CharacteristicsUtama Hadiputra SurbaktiNo ratings yet
- American Jewish Yearbook 1993 SingerDocument58 pagesAmerican Jewish Yearbook 1993 SingerJohn FuerstNo ratings yet
- The Great Migration and African-American Genomic Diversity: Simon - Gravel@mcgill - CaDocument27 pagesThe Great Migration and African-American Genomic Diversity: Simon - Gravel@mcgill - CaJohn FuerstNo ratings yet
- Reasoning Test Users Guide 2018Document29 pagesReasoning Test Users Guide 2018John FuerstNo ratings yet
- Weiss Saklofske PDFDocument9 pagesWeiss Saklofske PDFJohn FuerstNo ratings yet
- Student Score DistributionsDocument3 pagesStudent Score DistributionsJohn FuerstNo ratings yet
- Biogeographic Ancestry, Cognitive Ability and Socioeconomic OutcomesDocument26 pagesBiogeographic Ancestry, Cognitive Ability and Socioeconomic OutcomesJohn FuerstNo ratings yet
- Driver Test Scores For 2017-18 by EthnicityDocument1 pageDriver Test Scores For 2017-18 by EthnicityJohn FuerstNo ratings yet
- Global Ancestry and Cognitive AbilityDocument30 pagesGlobal Ancestry and Cognitive AbilityJohn FuerstNo ratings yet
- NCBI Infantil Spasme PDFDocument5 pagesNCBI Infantil Spasme PDFRiri Dwi AnggaraniNo ratings yet
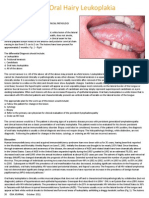
- Hairy LeukoplakiaDocument2 pagesHairy LeukoplakiaEka Septariana PuspaNo ratings yet
- Pages From (Ab) Normal Psychology 5th Ed. (Intro TXT) - S. Nolen-Hoeksema (McGraw-Hill, 2011) BBS CHAP 2Document73 pagesPages From (Ab) Normal Psychology 5th Ed. (Intro TXT) - S. Nolen-Hoeksema (McGraw-Hill, 2011) BBS CHAP 2Angel LacandulaNo ratings yet
- BiotransformationDocument27 pagesBiotransformationDemy ClementeNo ratings yet
- What Causes Glomerular DiseaseDocument5 pagesWhat Causes Glomerular DiseaseAswin AgusNo ratings yet
- The Biological Foundation of Behavior: Nervous, Endocrine and Immune SystemsDocument11 pagesThe Biological Foundation of Behavior: Nervous, Endocrine and Immune Systemsaudree d. aldayNo ratings yet
- How To Read A Next-Generation Sequencing Report-What Oncologists Need To KnowDocument9 pagesHow To Read A Next-Generation Sequencing Report-What Oncologists Need To KnowsamuelNo ratings yet
- Chapter 18 Endocrine SystemDocument40 pagesChapter 18 Endocrine SystemlolasparkleNo ratings yet
- Chapter 3 Recombinant DNA TechnologyDocument39 pagesChapter 3 Recombinant DNA TechnologyLovryan AmilingNo ratings yet
- Cardiovascular Blood VesselsDocument12 pagesCardiovascular Blood VesselswatuwaitingforNo ratings yet
- Dr. Ravi MalikDocument16 pagesDr. Ravi Malikkaabhay8866No ratings yet
- Entire BookDocument387 pagesEntire BookHillarie MeenachNo ratings yet
- Urinary System: G R O U P 4 - B S N 1 - 5Document59 pagesUrinary System: G R O U P 4 - B S N 1 - 5Love Jovellanos ÜNo ratings yet
- HCC HarrisonDocument5 pagesHCC HarrisonJesly CharliesNo ratings yet
- Popgen 2Document23 pagesPopgen 2teryNo ratings yet
- Atcm Journal 232 Sept 2016Document60 pagesAtcm Journal 232 Sept 2016Danilo NegrãoNo ratings yet
- Mirey - 2021 - Genotoxic Shift After Toxicants Exposure Modeling and Classification With A Multi-Endpoint ApproachDocument20 pagesMirey - 2021 - Genotoxic Shift After Toxicants Exposure Modeling and Classification With A Multi-Endpoint Approachyannick brunatoNo ratings yet
- Kami Export - 9.4 - UsingTheHardyWeinbergEquation - Worksheet Danielle Ngambia TchetmiDocument3 pagesKami Export - 9.4 - UsingTheHardyWeinbergEquation - Worksheet Danielle Ngambia TchetmijongambiaNo ratings yet
- Anatomical and Pathophysiological Classification of Congenital Heart DiseaseDocument16 pagesAnatomical and Pathophysiological Classification of Congenital Heart DiseasePietro PensoNo ratings yet
- Cerebral Circulation Anatomy and PhysiologyDocument5 pagesCerebral Circulation Anatomy and PhysiologyMayrjun Lo JacosalemNo ratings yet
- Aspek Laboratorium Kelainan HemostasisDocument48 pagesAspek Laboratorium Kelainan HemostasisriskypiradeNo ratings yet
- Assessment of Cadmium Tolerance and Biosorptive Potential of GCFSD01 Isolated From Cadmium Contaminated SoilDocument8 pagesAssessment of Cadmium Tolerance and Biosorptive Potential of GCFSD01 Isolated From Cadmium Contaminated SoilPaola PaezNo ratings yet
- Chapter 4 Experimental Embryology AnswersDocument2 pagesChapter 4 Experimental Embryology Answersbiblioteca10.uni.patxiNo ratings yet
- Chapter 28 Morphologic and Distributive Leukocyte DisordersDocument4 pagesChapter 28 Morphologic and Distributive Leukocyte DisordersAngeli CrisolNo ratings yet
- Hemoglobin Fraction in Thalassemia Patients Using Capillary ElectrophoresisDocument9 pagesHemoglobin Fraction in Thalassemia Patients Using Capillary ElectrophoresisYosua Butar ButarNo ratings yet
- 00713876fdgdfh41 CP PDFDocument1 page00713876fdgdfh41 CP PDFElisa SalakayNo ratings yet
- InfertilityDocument10 pagesInfertilityits_piks7256No ratings yet
- NemonicsDocument5 pagesNemonicsDsdNo ratings yet
- Commercial InvoiceDocument3 pagesCommercial InvoiceLJ RavenNo ratings yet
- July 28Document8 pagesJuly 28thestudentageNo ratings yet
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (18)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (78)
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 4 out of 5 stars4/5 (5)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (403)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (42)
- 12 Rules for Life by Jordan B. Peterson - Book Summary: An Antidote to ChaosFrom Everand12 Rules for Life by Jordan B. Peterson - Book Summary: An Antidote to ChaosRating: 4.5 out of 5 stars4.5/5 (207)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeNo ratings yet
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (169)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaFrom EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaRating: 4.5 out of 5 stars4.5/5 (266)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryFrom EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryRating: 4 out of 5 stars4/5 (44)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (328)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- The Happiness Trap: How to Stop Struggling and Start LivingFrom EverandThe Happiness Trap: How to Stop Struggling and Start LivingRating: 4 out of 5 stars4/5 (1)
- Algorithms to Live By: The Computer Science of Human DecisionsFrom EverandAlgorithms to Live By: The Computer Science of Human DecisionsRating: 4.5 out of 5 stars4.5/5 (722)
- Summary: How to Be an Adult in Relationships: The Five Keys to Mindful Loving by David Richo: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: How to Be an Adult in Relationships: The Five Keys to Mindful Loving by David Richo: Key Takeaways, Summary & Analysis IncludedRating: 4 out of 5 stars4/5 (11)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Secure Love: Create a Relationship That Lasts a LifetimeFrom EverandSecure Love: Create a Relationship That Lasts a LifetimeRating: 5 out of 5 stars5/5 (18)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisFrom EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (1)