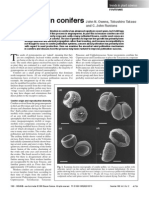
You might also like
- The Behavior of the Honey Bee in Pollen CollectionFrom EverandThe Behavior of the Honey Bee in Pollen CollectionNo ratings yet
- Glinos Cocucci 2011Document11 pagesGlinos Cocucci 2011Juan David Rodriguez HurtadoNo ratings yet
- Pollination: The Enduring Relationship between Plant and PollinatorFrom EverandPollination: The Enduring Relationship between Plant and PollinatorNo ratings yet
- Owens Et Al. 1998. Pollination in Conifers PDFDocument7 pagesOwens Et Al. 1998. Pollination in Conifers PDFJeffrey KellyNo ratings yet
- Ephedra Americana: The University of Chicago PressDocument15 pagesEphedra Americana: The University of Chicago Press1254789No ratings yet
- Anatomy Phalaenopsis AmabilisDocument8 pagesAnatomy Phalaenopsis AmabilisGenee Guevara MoonNo ratings yet
- Bird Visitation To A High Andean AnthuriumDocument6 pagesBird Visitation To A High Andean AnthuriumCamilo Barrios CardozoNo ratings yet
- Rutiliocarpun Caracolito InglésDocument7 pagesRutiliocarpun Caracolito InglésiNGRID VARGAS ROMERONo ratings yet
- TMP 3 D64Document9 pagesTMP 3 D64FrontiersNo ratings yet
- Krenn 2005Document40 pagesKrenn 2005Aziz Ait AkkaNo ratings yet
- AngiospermDocument5 pagesAngiospermAbdul BasitNo ratings yet
- 1 s2.0 S0254629910001894 Main PDFDocument12 pages1 s2.0 S0254629910001894 Main PDFcamiloNo ratings yet
- Origin AngiospermsDocument5 pagesOrigin Angiospermsmustafa md100% (1)
- American J of Botany - 2005 - Zhang - Predicting Mating Patterns From Pollination Syndromes The Case of SapromyiophilyDocument8 pagesAmerican J of Botany - 2005 - Zhang - Predicting Mating Patterns From Pollination Syndromes The Case of Sapromyiophily罗依婷No ratings yet
- Tschapka - Dressler. 2002. ChiropterophyliDocument15 pagesTschapka - Dressler. 2002. ChiropterophyliJ E Fernando CernaNo ratings yet
- Phylogenetic Analysis of Eurytominae (Chalcidoidea: Eurytomidae) Based On Morphological CharactersDocument70 pagesPhylogenetic Analysis of Eurytominae (Chalcidoidea: Eurytomidae) Based On Morphological CharactersAna MaríaNo ratings yet
- Reproductive Biology of Erythrina Falcata (Fabaceae: Papilionoideae) 1Document11 pagesReproductive Biology of Erythrina Falcata (Fabaceae: Papilionoideae) 1michaelNo ratings yet
- Asociación Planta OrmigaDocument4 pagesAsociación Planta OrmigaDaniel Aparicio HilaresNo ratings yet
- Investigation On Seedling Morphology in Taxonomic StudiesDocument9 pagesInvestigation On Seedling Morphology in Taxonomic StudiesCarolineVasconcelosNo ratings yet
- Floral Rewards and Pollination in Cytiseae (Fabaceae) : Plant Systematics and Evolution May 2003Document12 pagesFloral Rewards and Pollination in Cytiseae (Fabaceae) : Plant Systematics and Evolution May 2003AngelaNo ratings yet
- APOCINACEAEDocument13 pagesAPOCINACEAEVenya Nabila VirdausyNo ratings yet
- Tobe FloralMorphologyEmbryology 1993Document7 pagesTobe FloralMorphologyEmbryology 1993Pancho FarroñayNo ratings yet
- Evolutionary Disruption in The Pollination System of Vanilla OrchidaceaeDocument11 pagesEvolutionary Disruption in The Pollination System of Vanilla Orchidaceaedennis.tanNo ratings yet
- The Ant-Pollination System of Cytinus Hypocistis (Cytinaceae), A Mediterranean Root HoloparasiteDocument11 pagesThe Ant-Pollination System of Cytinus Hypocistis (Cytinaceae), A Mediterranean Root HoloparasiterachiiidaNo ratings yet
- Notes On The Floral Biology of NymphaeaDocument4 pagesNotes On The Floral Biology of NymphaeaLeandrorvNo ratings yet
- MCP 049Document11 pagesMCP 049rachiiidaNo ratings yet
- Endress 2008 - Perianth Biology in The Basal Grade of Extant AngiospermsDocument20 pagesEndress 2008 - Perianth Biology in The Basal Grade of Extant AngiospermsMárcio BazanteNo ratings yet
- Angiosperms Origin and EvolutionDocument25 pagesAngiosperms Origin and EvolutionShah RafiqNo ratings yet
- Synthesis Paper: What Is FlowerDocument5 pagesSynthesis Paper: What Is Flowercristhel joyNo ratings yet
- Pteridophyte (Introductio, Reproduction Etc.)Document15 pagesPteridophyte (Introductio, Reproduction Etc.)Joy GhoshNo ratings yet
- A Scanning Electron Microscopy Study of Pollen GrainsDocument10 pagesA Scanning Electron Microscopy Study of Pollen GrainsIJAR JOURNALNo ratings yet
- British Ecological Society Journal of Ecology: This Content Downloaded From 200.24.16.65 On Sat, 08 Jun 2019 16:36:43 UTCDocument13 pagesBritish Ecological Society Journal of Ecology: This Content Downloaded From 200.24.16.65 On Sat, 08 Jun 2019 16:36:43 UTCJESSICA HERRERA SANCHEZNo ratings yet
- Angiosperms - Origin Amd EvolutionDocument24 pagesAngiosperms - Origin Amd EvolutionYogesh SharmaNo ratings yet
- Topic: Angiosperm: DescriptionDocument12 pagesTopic: Angiosperm: DescriptionVINDHYA SHANKERNo ratings yet
- Aglaophyton: Temporal Range: Early DevonianDocument3 pagesAglaophyton: Temporal Range: Early Devonianx456456456xNo ratings yet
- Nectar ReviewDocument16 pagesNectar ReviewEliezer BecziNo ratings yet
- Gonzlezetal Boletin SABDocument11 pagesGonzlezetal Boletin SABANDRÉS DÍAZ IBÁÑEZNo ratings yet
- Kingdom Plantae CompleteDocument21 pagesKingdom Plantae Completegabrielluis08No ratings yet
- Studies On Biology and Reproduction of Butterflies (Family: Papilionidae) in Nilgiris Hills, Southern Western Ghats, IndiaDocument11 pagesStudies On Biology and Reproduction of Butterflies (Family: Papilionidae) in Nilgiris Hills, Southern Western Ghats, IndiaEman SamirNo ratings yet
- Preferensi Kupu-Kupu Familia Papilionidae Dan Pieridae Pada Tumbuhan Di Wisata Air Terjun Coban Rais Kota Batu, Jawa TimurDocument7 pagesPreferensi Kupu-Kupu Familia Papilionidae Dan Pieridae Pada Tumbuhan Di Wisata Air Terjun Coban Rais Kota Batu, Jawa Timurفواش فلاسفNo ratings yet
- American J of Botany - 2009 - Taylor - Seed Ferns From The Late Paleozoic and Mesozoic Any Angiosperm Ancestors LurkingDocument15 pagesAmerican J of Botany - 2009 - Taylor - Seed Ferns From The Late Paleozoic and Mesozoic Any Angiosperm Ancestors LurkingbsdunleyNo ratings yet
- Orchid Pollination Biology: Haleigh Ray and Wagner VendrameDocument6 pagesOrchid Pollination Biology: Haleigh Ray and Wagner VendrameSue OrangesNo ratings yet
- Flower Visitor Interaction and Fruit Production of Grewia Asiatica LDocument7 pagesFlower Visitor Interaction and Fruit Production of Grewia Asiatica LBotany with HyderAliNo ratings yet
- Community Structure of Sporulating Fungi On Decaying Litters of Shorea SPPDocument8 pagesCommunity Structure of Sporulating Fungi On Decaying Litters of Shorea SPPSitifatimahNo ratings yet
- Morphological Varia Tions of Fertile Spik e in Helminthostachys Zeylanica (L.) HookDocument5 pagesMorphological Varia Tions of Fertile Spik e in Helminthostachys Zeylanica (L.) HookSandip Dev ChaudhuriNo ratings yet
- Flores Generalistas, Biodiversidad y Florivoria, Implicaciones para El Origen de Las AngiospermasDocument6 pagesFlores Generalistas, Biodiversidad y Florivoria, Implicaciones para El Origen de Las AngiospermasRobinNo ratings yet
- 1612 FullDocument18 pages1612 Fulljalvarez7693No ratings yet
- Anatomical Patterns of Aerenchyma in Aquatic Plants 2008Document12 pagesAnatomical Patterns of Aerenchyma in Aquatic Plants 2008Gilberto Aleman SancheschulzNo ratings yet
- Gilbert 1994Document7 pagesGilbert 1994Jhon W. MuñuicoNo ratings yet
- Orchids of NepalDocument145 pagesOrchids of Nepalshinkaron88100% (1)
- Ecology 7Document16 pagesEcology 7JESSICA HERRERA SANCHEZNo ratings yet
- GraciaDocument24 pagesGraciaaini salimNo ratings yet
- 1 1999 FlacourtiaceaeDocument16 pages1 1999 FlacourtiaceaeJuan David Rodriguez HurtadoNo ratings yet
- 12th Class Biology Ncert Chap-2Document23 pages12th Class Biology Ncert Chap-2Prathyusha MannemNo ratings yet
- Anthoceros - General FeaturesDocument22 pagesAnthoceros - General FeaturesTanishka Aggarwal100% (1)
- (Pollen Morphology of Neotropical Species of Podostemum (Malpighiales: Podostemaceae) )Document9 pages(Pollen Morphology of Neotropical Species of Podostemum (Malpighiales: Podostemaceae) )NilsonNo ratings yet
- Inflorescence CyperaceaeInflorDocument21 pagesInflorescence CyperaceaeInflorGhulamJelaniNo ratings yet
- OrchidsDocument12 pagesOrchidsjomeraranaNo ratings yet
- Pollen Morphological Studies of Selected Plants of Asteraceae FamilyDocument8 pagesPollen Morphological Studies of Selected Plants of Asteraceae FamilyIJRASETPublicationsNo ratings yet
- Angiosperm Derived CharacteristicsDocument7 pagesAngiosperm Derived CharacteristicsBreen DoteNo ratings yet
- Pollination Biologi of Hydrangea MacrophyllaDocument5 pagesPollination Biologi of Hydrangea MacrophyllakhairulNo ratings yet
- Pollination WikipediaDocument12 pagesPollination WikipediaAnirban MallickNo ratings yet
- Pollination - Biology Science Fair Project IdeasDocument10 pagesPollination - Biology Science Fair Project IdeasSwaraj KhedekarNo ratings yet
- Sexual Reproduction in Flowering Plants: 2.1 F - A F O ADocument23 pagesSexual Reproduction in Flowering Plants: 2.1 F - A F O AVikram SkNo ratings yet
- Reproduction in Flowering Plants: "Imperfect or Incomplete Flowers"Document7 pagesReproduction in Flowering Plants: "Imperfect or Incomplete Flowers"Jann LarNo ratings yet
- Neet Wizz Bio Class 12Document89 pagesNeet Wizz Bio Class 12Aarthi T. UNo ratings yet
- Reproduction in Flowering Plants PrintDocument10 pagesReproduction in Flowering Plants PrintSarada KasyapNo ratings yet
- Interactive Key Notes On Sexual Reproduction in Flowering PlantsDocument16 pagesInteractive Key Notes On Sexual Reproduction in Flowering PlantsnestNo ratings yet
- Skema Trial Biologi MRSM K2Document13 pagesSkema Trial Biologi MRSM K2sh. ainin sofiyaNo ratings yet
- Botany I (EM) BLM 2021-22Document67 pagesBotany I (EM) BLM 2021-22Ram CharanNo ratings yet
- UNIT 2 Notes PDFDocument88 pagesUNIT 2 Notes PDFAjNo ratings yet
- Worksheet ReproductionDocument8 pagesWorksheet ReproductionStefani KavangoNo ratings yet
- Sexual Repro PlantDocument29 pagesSexual Repro PlantAshwani PathakNo ratings yet
- Reproduction PPT 3 ANGIOSPERMSDocument76 pagesReproduction PPT 3 ANGIOSPERMSlisanames.23No ratings yet
- Biology SPM 2020 Model Answers For Predicted QuestionsDocument30 pagesBiology SPM 2020 Model Answers For Predicted Questionskumarantuition100% (1)
- Final4 Merged New 4Document95 pagesFinal4 Merged New 4rohanNo ratings yet
- Pollen GerminationDocument6 pagesPollen GerminationJustyNo ratings yet
- Topic 3 BioDocument39 pagesTopic 3 BioTsukiNo ratings yet
- Plant Reproduction: L1 - FlowersDocument56 pagesPlant Reproduction: L1 - FlowersMaha NaserNo ratings yet
- Detailed Lesson Plan in BiologyDocument23 pagesDetailed Lesson Plan in BiologyBernard Ortinero100% (5)
- Pollination in Sunflower and LegumDocument18 pagesPollination in Sunflower and Legumkanha kumarNo ratings yet
- Reproduction in PlantsDocument12 pagesReproduction in PlantsAnand Philip PrasadNo ratings yet
- XII BiologyDocument114 pagesXII BiologyGuruKPO100% (1)
- Bio CH6 F5 StudywithadminDocument7 pagesBio CH6 F5 StudywithadminRathi MalarNo ratings yet
- Pollination & Fruit Development in TomatoesDocument4 pagesPollination & Fruit Development in Tomatoeshung nguyen vietNo ratings yet
- Biology Multiple Choice QuestionDocument20 pagesBiology Multiple Choice QuestionNalina BalasingamNo ratings yet
- Reproduction in PlantsDocument68 pagesReproduction in PlantsZainurain Idrissi SchoolNo ratings yet
- Biology Project: A Cbse English Medium Co-Educational Senior Secondary School SESSION: 2021-2022Document10 pagesBiology Project: A Cbse English Medium Co-Educational Senior Secondary School SESSION: 2021-2022Omar Abdullah100% (1)
- Block-06 GymnospermsDocument82 pagesBlock-06 Gymnospermstamil selviNo ratings yet
- Sexual Reproduction in Flowering PlantsDocument66 pagesSexual Reproduction in Flowering Plantsshobitgup100% (1)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (6)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (516)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)
- Lymph & Longevity: The Untapped Secret to HealthFrom EverandLymph & Longevity: The Untapped Secret to HealthRating: 4.5 out of 5 stars4.5/5 (13)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (4)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Change Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessFrom EverandChange Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessRating: 4 out of 5 stars4/5 (18)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)