You might also like
- Immuno 3aprilDocument84 pagesImmuno 3aprilAlkaNo ratings yet
- Tcell Maturation and Selection - ReportDocument10 pagesTcell Maturation and Selection - ReportShivendra VermaNo ratings yet
- CMI EnglishDocument5 pagesCMI EnglishAndika PesakNo ratings yet
- T Cells & B Cells: 6.1 - Antigen Processing and PresentationDocument6 pagesT Cells & B Cells: 6.1 - Antigen Processing and PresentationAmanda AmiraultNo ratings yet
- Maduracion y Activacion de Celulas T Clase 1 y 2Document4 pagesMaduracion y Activacion de Celulas T Clase 1 y 2EDGAR FUENMAYOR PIRELANo ratings yet
- Clonal SelectionDocument65 pagesClonal SelectionInjamam ul hossainNo ratings yet
- Unit 8 T CellDocument33 pagesUnit 8 T CellamirahaliNo ratings yet
- Parcial 3 Inmunología T Cell DevelopmentDocument4 pagesParcial 3 Inmunología T Cell Developmentjuan mondaNo ratings yet
- TCR and T Lymphocyte Development in ThymusDocument39 pagesTCR and T Lymphocyte Development in ThymusWawan WaeNo ratings yet
- Chapter 15Document9 pagesChapter 15fairy_topia_2007No ratings yet
- Immunopathology: Normal Immune ResponseDocument13 pagesImmunopathology: Normal Immune ResponseChinedu H. DuruNo ratings yet
- Lecture - T Cell DevelopmentDocument29 pagesLecture - T Cell Developmentrockyrawat01012003No ratings yet
- Crossroads of Extrathymic Lymphocytes Maturation PathwaysDocument5 pagesCrossroads of Extrathymic Lymphocytes Maturation PathwaysIvan BubanovicNo ratings yet
- T-Cell Development in The ThymusDocument1 pageT-Cell Development in The Thymusfysi mackNo ratings yet
- Cell-Mediated Immunity: A. Antigen - Specific CellsDocument7 pagesCell-Mediated Immunity: A. Antigen - Specific CellsandindlNo ratings yet
- Cancer Immunotherapy 2020Document76 pagesCancer Immunotherapy 2020stupid.graphsNo ratings yet
- Haematology: Lecture NotesDocument4 pagesHaematology: Lecture NotesjonasNo ratings yet
- What Are The Main Cells of The Immune System?Document5 pagesWhat Are The Main Cells of The Immune System?Idrissa John Sebeh ContehNo ratings yet
- Sam CD Rom - ImmunologyDocument111 pagesSam CD Rom - ImmunologyDr. Muha. Hasan Mahbub-Ur-RahmanNo ratings yet
- Bahan Kuliah Id-1-2020Document63 pagesBahan Kuliah Id-1-2020Andi SaputraNo ratings yet
- 5 Adaptive ImmunityDocument35 pages5 Adaptive Immunityozerbilge24No ratings yet
- Paracortex Where Naïve Helper T Cells & Cytotoxic T Cells Are LocatedDocument4 pagesParacortex Where Naïve Helper T Cells & Cytotoxic T Cells Are LocatedNabeel ShahzadNo ratings yet
- Immnunology Notebook Chapter One: Innate ImmunityDocument45 pagesImmnunology Notebook Chapter One: Innate ImmunityJavier Alejandro Daza GalvánNo ratings yet
- محاضرة دكتورة ايمانDocument9 pagesمحاضرة دكتورة ايمانMsikvNo ratings yet
- Cellular MechanismsDocument20 pagesCellular MechanismsPriyanshu BlazeNo ratings yet
- C-12-Tcell and Bcell ActivationDocument43 pagesC-12-Tcell and Bcell Activationlinnet17No ratings yet
- T Cell Maturation, Activation and DifferentiationDocument65 pagesT Cell Maturation, Activation and DifferentiationChanDan RjNo ratings yet
- Development of Imune CellsDocument28 pagesDevelopment of Imune CellsAkss ShauryaNo ratings yet
- T CellDocument36 pagesT CellSurja DasNo ratings yet
- Lecture 11Document31 pagesLecture 11Binita SedhaiNo ratings yet
- Cell-Mediated Immune Responses8Document26 pagesCell-Mediated Immune Responses8CORYNEBACTERIUMNo ratings yet
- Lecture 8: Adaptive ImmunityDocument3 pagesLecture 8: Adaptive ImmunityMohit ModyNo ratings yet
- T-Cell Dependent B - Cell DependentDocument29 pagesT-Cell Dependent B - Cell DependentGayathri deviNo ratings yet
- Cell Mediated ImmunityDocument22 pagesCell Mediated ImmunityDrJoko SulistyoNo ratings yet
- T Cell Subsets and T Cell-Mediated Immunity: Femke Broere, Sergei G. Apasov, Michail V. Sitkovsky and Willem Van EdenDocument14 pagesT Cell Subsets and T Cell-Mediated Immunity: Femke Broere, Sergei G. Apasov, Michail V. Sitkovsky and Willem Van EdenDavion StewartNo ratings yet
- TI1 and TI2Document4 pagesTI1 and TI2sultan khabeebNo ratings yet
- Chapter 9 T-Cell Maturation, ActivationDocument65 pagesChapter 9 T-Cell Maturation, ActivationTofikNo ratings yet
- 7 Chapter 7 T-Cell Maturation, Activation, and DifferentiationDocument34 pages7 Chapter 7 T-Cell Maturation, Activation, and DifferentiationMekuriya BeregaNo ratings yet
- Cell Surface Markers of T-Cells, B-Cells andDocument6 pagesCell Surface Markers of T-Cells, B-Cells andUdaya KumarNo ratings yet
- The Immune System & Lymphoid Organs: Bulseco, Carino, Dinglasan, SalanguitDocument88 pagesThe Immune System & Lymphoid Organs: Bulseco, Carino, Dinglasan, SalanguitVanessa CarinoNo ratings yet
- T Lymphocytes: Cells in FocusDocument5 pagesT Lymphocytes: Cells in FocusIvan VazquezNo ratings yet
- 05 - Adaptive Immunity (T Cell Receptors)Document21 pages05 - Adaptive Immunity (T Cell Receptors)star lightNo ratings yet
- Presented By:: Rehnuma K. GhoriDocument28 pagesPresented By:: Rehnuma K. GhoriavaniNo ratings yet
- Fundamentals of Pathology The Adaptive Immune Response T CellsDocument41 pagesFundamentals of Pathology The Adaptive Immune Response T CellsOsman MNo ratings yet
- Immune SystemDocument32 pagesImmune Systemanupama menonNo ratings yet
- Immuno PathologyDocument28 pagesImmuno PathologyIdrissa John Sebeh ContehNo ratings yet
- Wiki T CellDocument12 pagesWiki T Cellالولد الخطير تابعونيNo ratings yet
- B Cell and T Cell DevelopmentDocument2 pagesB Cell and T Cell Developmentzeynep sevinçNo ratings yet
- 2022 Article 210Document13 pages2022 Article 210uttamNo ratings yet
- Lecture - TCRDocument9 pagesLecture - TCRrockyrawat01012003No ratings yet
- Intro To ImmunologyDocument33 pagesIntro To ImmunologyandreaNo ratings yet
- T-Cells-1Document156 pagesT-Cells-1pilot abdi baariNo ratings yet
- T CellDocument19 pagesT CellSachin AnandNo ratings yet
- T Cell - WikipediaDocument65 pagesT Cell - WikipediaPowell KitagwaNo ratings yet
- Cell-Mediated ImmunityDocument52 pagesCell-Mediated ImmunityAhmed AlosaiqueNo ratings yet
- MHC and Disease Susceptibility (Type 1 Diabetes As Example)Document8 pagesMHC and Disease Susceptibility (Type 1 Diabetes As Example)FATbovenNo ratings yet
- Principles of Transplantation SHDocument93 pagesPrinciples of Transplantation SHqadiraftabNo ratings yet
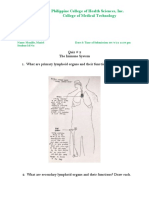
- Philippine College of Health Sciences, Inc. College of Medical TechnologyDocument6 pagesPhilippine College of Health Sciences, Inc. College of Medical TechnologyMariel AbellezaNo ratings yet
- Kaufmann 1996Document8 pagesKaufmann 1996Yasmin Antunes de Abreu Dos SantosNo ratings yet
- Fast Facts: CAR T-Cell Therapy: Insight into current and future applicationsFrom EverandFast Facts: CAR T-Cell Therapy: Insight into current and future applicationsNo ratings yet
- Immuno TransplantationDocument31 pagesImmuno Transplantationalka mehraNo ratings yet
- ToleranceDocument39 pagesTolerancealka mehraNo ratings yet
- Immuno SemVI-Lec.2 StudentsDocument6 pagesImmuno SemVI-Lec.2 Studentsalka mehraNo ratings yet
- Crick PrsentationDocument28 pagesCrick Prsentationalka mehraNo ratings yet
- Supre Re Soul Tion ConceptsDocument5 pagesSupre Re Soul Tion Conceptsalka mehraNo ratings yet
- Indo US Workshop - Mass Spec and MetabolomicsDocument22 pagesIndo US Workshop - Mass Spec and Metabolomicsalka mehraNo ratings yet
- BScthesis On MDDocument46 pagesBScthesis On MDalka mehraNo ratings yet
- Lecture2 RDT 1.02.2023 PDocument20 pagesLecture2 RDT 1.02.2023 Palka mehraNo ratings yet
- VCFv4.2 FormatDocument29 pagesVCFv4.2 Formatalka mehraNo ratings yet
- HEMA-1 Up To PG 30.-1-31Document31 pagesHEMA-1 Up To PG 30.-1-31Angela ReyesNo ratings yet
- Switching Homes: How Cancer Moves To Bone: Molecular SciencesDocument25 pagesSwitching Homes: How Cancer Moves To Bone: Molecular SciencesNashely GarcíaNo ratings yet
- Hematology 1 Mtap1Document70 pagesHematology 1 Mtap1Cyril Llego ManuelNo ratings yet
- Introduction To Stem Cells: Speaker - Maj Mohd Anas Sheikh Guide - LT Col AK BiswasDocument38 pagesIntroduction To Stem Cells: Speaker - Maj Mohd Anas Sheikh Guide - LT Col AK BiswasMohd Anas SheikhNo ratings yet
- GeneticsDocument246 pagesGeneticsVinita Saroha100% (9)
- Final Thesis (1) - Merged - CompressedDocument195 pagesFinal Thesis (1) - Merged - CompressedGhassan KhudhairNo ratings yet
- Hematology Book For PDF 1304281501 Phpapp02 PDFDocument260 pagesHematology Book For PDF 1304281501 Phpapp02 PDFAndrea Kocsis100% (1)
- Life Flows Because of You..Document40 pagesLife Flows Because of You..Klemen JamnikNo ratings yet
- Hematopoiesis PDFDocument10 pagesHematopoiesis PDFJezzah Mae CañeteNo ratings yet
- Hepatic Stem Cells Methods and Protocols - Naoki Tanimizu - 2019 PDFDocument242 pagesHepatic Stem Cells Methods and Protocols - Naoki Tanimizu - 2019 PDFTristan PhilipsNo ratings yet
- B Cells in Immunity and Tolerance - BookDocument190 pagesB Cells in Immunity and Tolerance - BookJuan FranciscoNo ratings yet
- (Visvader Et Al., 2016) Tissue-Specific Designs of Stem Cell HierarchiesDocument7 pages(Visvader Et Al., 2016) Tissue-Specific Designs of Stem Cell HierarchiesJulio dR AltavasNo ratings yet
- Roles of Vitamins in Stem Cells: Cellular and Molecular Life Sciences May 2020Document22 pagesRoles of Vitamins in Stem Cells: Cellular and Molecular Life Sciences May 2020Chanda DajaniNo ratings yet
- Humanized Mice For The Study of Immuno-Oncology: ReviewDocument16 pagesHumanized Mice For The Study of Immuno-Oncology: ReviewMaria Camila MejíaNo ratings yet
- Jungzelleneffekt Autophagy Dev146506.fullDocument14 pagesJungzelleneffekt Autophagy Dev146506.fullmillywanillyNo ratings yet
- After Effects of COVID 19 and Repair of LungsDocument13 pagesAfter Effects of COVID 19 and Repair of LungsAthenaeum Scientific PublishersNo ratings yet
- Statnotes PDFDocument300 pagesStatnotes PDFSivanantham KrishnamoorthiNo ratings yet
- Haematology: Lecture NotesDocument15 pagesHaematology: Lecture Notesamanuel melakuNo ratings yet
- Primary and Secondary Lymphoid Organs - Aditi SinghDocument50 pagesPrimary and Secondary Lymphoid Organs - Aditi SinghEunice PalloganNo ratings yet
- Development and Maintenance of Hematopoietic Tissues: Colony-Forming Units (Cfus)Document13 pagesDevelopment and Maintenance of Hematopoietic Tissues: Colony-Forming Units (Cfus)Tyler KingNo ratings yet
- CV Kajaste Novembre2023Document8 pagesCV Kajaste Novembre2023powersplyNo ratings yet
- Small Intestinal Stem Cell Markers: Robert K. Montgomery and David T. BreaultDocument7 pagesSmall Intestinal Stem Cell Markers: Robert K. Montgomery and David T. BreaultAbdullahNo ratings yet
- Article Text 75576 1 10 20210527Document10 pagesArticle Text 75576 1 10 20210527Nimbus MultiversesNo ratings yet
- Cytoplasmic Labile Iron Accumulates in Aging Stem Cells Perturbing A Key Rheostat For Identity ControlDocument71 pagesCytoplasmic Labile Iron Accumulates in Aging Stem Cells Perturbing A Key Rheostat For Identity ControlValen EstevezNo ratings yet
- Haemnatopoiesis and CMLDocument8 pagesHaemnatopoiesis and CMLKe XuNo ratings yet
- Serum-Free Medium and ApplicationsDocument4 pagesSerum-Free Medium and Applicationsharshita sharmaNo ratings yet
- OsteoinmunologíaDocument19 pagesOsteoinmunologíaKate RodasNo ratings yet
- Bone Marrow Evaluation For Pediatric PatientsDocument7 pagesBone Marrow Evaluation For Pediatric PatientssylvetriNo ratings yet
- Avian Histophatology, 4th Edition (VetBooks - Ir) PDFDocument664 pagesAvian Histophatology, 4th Edition (VetBooks - Ir) PDFAnn Tyshkivska100% (4)
- Annual Report 06-07englishDocument277 pagesAnnual Report 06-07englisharvindftNo ratings yet