You might also like
- Caecilians Venom GlandsDocument18 pagesCaecilians Venom GlandsAngel AyalaNo ratings yet
- Early Development of Betta Splendens Under Stereomicroscopy and Scanning Electron MicrosDocument11 pagesEarly Development of Betta Splendens Under Stereomicroscopy and Scanning Electron Micros0 9No ratings yet
- Research Highlights: An Open and Shut CaseDocument2 pagesResearch Highlights: An Open and Shut Caseapi-26828680No ratings yet
- AMGH V52i4 2Document71 pagesAMGH V52i4 2Valentina Renza PinoNo ratings yet
- Tilapia WelfareDocument17 pagesTilapia Welfareমুহাম্মাদ রিদুয়ান পাশাNo ratings yet
- Diversity and Biotechnological Potential of Culturable Bacteria From Brazilian Mangrove SedimentDocument8 pagesDiversity and Biotechnological Potential of Culturable Bacteria From Brazilian Mangrove SedimentMuhammad Wildan GunaryaNo ratings yet
- Enterocins Symptomatic For Bio Alternative in Caries ControlDocument7 pagesEnterocins Symptomatic For Bio Alternative in Caries ControlSSR-IIJLS JournalNo ratings yet
- ALaboratory Reportonthe Observed Parasitesinthe Gillsand Gutof Anabastestudineus Puyofrom Kabacan Cotabato PhilippinesDocument15 pagesALaboratory Reportonthe Observed Parasitesinthe Gillsand Gutof Anabastestudineus Puyofrom Kabacan Cotabato PhilippinesNicollo DadiavelliNo ratings yet
- Micro-Dissection of Enamel Organ From Mandibular Incisor of Rats Exposed To Environmental ToxicantsDocument10 pagesMicro-Dissection of Enamel Organ From Mandibular Incisor of Rats Exposed To Environmental ToxicantsNadira NurinNo ratings yet
- Final PaperDocument26 pagesFinal PaperGeriz Daniella VigoNo ratings yet
- 2018 - Valente Et Al - B Jararaca Accessory Venom Gland Ancillary Source of ToxinsDocument12 pages2018 - Valente Et Al - B Jararaca Accessory Venom Gland Ancillary Source of ToxinsFerenc TörökNo ratings yet
- Reseasch On GeolocationDocument3 pagesReseasch On Geolocationamarenderashish0802No ratings yet
- Associations Between Salivary Levels of Streptococcus Mutans, Streptococcus Sobrinus, Lactobacilli and Caries Experience in Kenyan AdolescentsDocument7 pagesAssociations Between Salivary Levels of Streptococcus Mutans, Streptococcus Sobrinus, Lactobacilli and Caries Experience in Kenyan AdolescentsgabyNo ratings yet
- Brain and Testis - More Alike Than Previously ThoughtDocument11 pagesBrain and Testis - More Alike Than Previously Thoughtrohit.jaruplaNo ratings yet
- Identi Cation of Proteins From Human Permanent Erupted Enamel Castiblanco GA, Rutishauser D, Ilag LL, Martignon S, CastDocument7 pagesIdenti Cation of Proteins From Human Permanent Erupted Enamel Castiblanco GA, Rutishauser D, Ilag LL, Martignon S, CastMoisés MotaNo ratings yet
- Parents With Periodontitis Drive The Early Acquisition of Dysbiotic Microbiomes in Their OffspringDocument16 pagesParents With Periodontitis Drive The Early Acquisition of Dysbiotic Microbiomes in Their Offspringjulijana.veipaNo ratings yet
- Biolog A Celular y Organism Os ModeloDocument75 pagesBiolog A Celular y Organism Os Modeloalanis fernanda haro benitesNo ratings yet
- Borgesetal 2020Document26 pagesBorgesetal 2020Daniel E. Zarcos LópezNo ratings yet
- Antibacterial Activity of Avocado Extracts (Persea Americana Mill.) Against Streptococcus AgalactiaeDocument8 pagesAntibacterial Activity of Avocado Extracts (Persea Americana Mill.) Against Streptococcus AgalactiaeAndrea KhudzeNo ratings yet
- ChickenPeyersPatchesJI1980 PDFDocument9 pagesChickenPeyersPatchesJI1980 PDFDulce AmorNo ratings yet
- Fundamentals of Practical Biology: For Senior Secondary Schools and CollegesFrom EverandFundamentals of Practical Biology: For Senior Secondary Schools and CollegesNo ratings yet
- Biomphalaria Embryotoxicity Test (BET) : 60 Years of Research Crossing Boundaries For Developing Standard ProtocolsDocument16 pagesBiomphalaria Embryotoxicity Test (BET) : 60 Years of Research Crossing Boundaries For Developing Standard ProtocolsJalison RêgoNo ratings yet
- GB 2011 12 7 406Document9 pagesGB 2011 12 7 406seebass srosrNo ratings yet
- Leche de CucarachaDocument12 pagesLeche de CucarachaMARCO MARTIN SALAZAR BALDEONNo ratings yet
- Enzymatic Profile of The Pacific WhiteDocument8 pagesEnzymatic Profile of The Pacific WhiteroraysideblancoNo ratings yet
- 2022 Dentaldefects Environmental EHP10208Document12 pages2022 Dentaldefects Environmental EHP10208Senussi IbtisamNo ratings yet
- Isolation and Identification of Proteolytic BacteriaDocument8 pagesIsolation and Identification of Proteolytic BacteriaMd. Samaul AlamNo ratings yet
- 2014 ReichardtismejDocument14 pages2014 ReichardtismejlalaaaaNo ratings yet
- Stefani 2012 Mesa BolivarDocument14 pagesStefani 2012 Mesa BolivarDyllandu.LNo ratings yet
- Week 5 Cellulose BreakdownDocument2 pagesWeek 5 Cellulose Breakdownapi-258192402No ratings yet
- Sundqvist, G. (1994) Taxonomy, Ecology, and Pathogenicity of The Root Canal Flora.Document9 pagesSundqvist, G. (1994) Taxonomy, Ecology, and Pathogenicity of The Root Canal Flora.Jaime BarraganNo ratings yet
- 2019 - Paiva Et Al - Osmophores of Caryocar Brasiliens A Particular Structure of The AndroeciumDocument12 pages2019 - Paiva Et Al - Osmophores of Caryocar Brasiliens A Particular Structure of The AndroeciumValeria CasiqueNo ratings yet
- Biogeografia - Arañas en ColombiaDocument49 pagesBiogeografia - Arañas en ColombiaLilia SierraNo ratings yet
- Electrical PolarizationDocument35 pagesElectrical PolarizationksdayaNo ratings yet
- Autism and The Gut: OutlookDocument2 pagesAutism and The Gut: OutlookLindo PulgosoNo ratings yet
- VISJ Issue 5 August 2021Document22 pagesVISJ Issue 5 August 2021Desirée VidalNo ratings yet
- Population Position Along The Fast Slow Life History Continuum Predicts Intraspecific Variation in Actuarial SenescenceDocument25 pagesPopulation Position Along The Fast Slow Life History Continuum Predicts Intraspecific Variation in Actuarial SenescenceDaniel GhiurcaNo ratings yet
- Heritable Variations of Pteris BiauritaDocument7 pagesHeritable Variations of Pteris Biauritacefaran159No ratings yet
- Clinical Parasitology Module 2: Macroscopic Evaluation of Stool & Intestinal NematodesDocument4 pagesClinical Parasitology Module 2: Macroscopic Evaluation of Stool & Intestinal NematodesJhona Mae CortesNo ratings yet
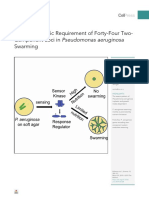
- Context-Specific Requirement of Forty-Four Two-Component Loci in Pseudomonas Aeruginosa SwarmingDocument27 pagesContext-Specific Requirement of Forty-Four Two-Component Loci in Pseudomonas Aeruginosa SwarmingmacNo ratings yet
- 52219-Texto Do Artigo-751375224346-1-10-20210426Document7 pages52219-Texto Do Artigo-751375224346-1-10-20210426Manu MorpheusNo ratings yet
- Ray Etal 2010 Microbes and Solids Management FINALDocument10 pagesRay Etal 2010 Microbes and Solids Management FINALBenjamin BarcelóNo ratings yet
- Velez Arellanoetal2014Document7 pagesVelez Arellanoetal2014Ingka SinaleNo ratings yet
- Fmicb 08 01226Document13 pagesFmicb 08 01226Gustavo MedinaNo ratings yet
- Morfologi Dan Biometri Spermatozoa Anoa (Bubalus SP.) Yang Diwarnai Dengan Pewarna William's Dan Eosin-NigrosinDocument8 pagesMorfologi Dan Biometri Spermatozoa Anoa (Bubalus SP.) Yang Diwarnai Dengan Pewarna William's Dan Eosin-NigrosinZiyad MalikNo ratings yet
- An Estimation of The Number of Cells in The Human BodyDocument12 pagesAn Estimation of The Number of Cells in The Human BodyrharaNo ratings yet
- Janiak Et Al 2020 Genetic Evidence of Widespread Variation in Ethanol Metabolism Among Mammals Revisiting The Myth ofDocument7 pagesJaniak Et Al 2020 Genetic Evidence of Widespread Variation in Ethanol Metabolism Among Mammals Revisiting The Myth ofAlvaro MakobiNo ratings yet
- Avocado 1Document8 pagesAvocado 1Feddy Micheal FeddyNo ratings yet
- DNA Barcoding of Freshwater Fishes From Lake Laut Tawar, Aceh Province, IndonesiaDocument10 pagesDNA Barcoding of Freshwater Fishes From Lake Laut Tawar, Aceh Province, IndonesiaQurratul 'AiniNo ratings yet
- N Micro Bio L 2016177Document5 pagesN Micro Bio L 2016177Luis Gustavo Calatayud GarcíaNo ratings yet
- A Prebiotic Matrix For Encapsulation of Probiotics: Physicochemical and Microbiological StudyDocument14 pagesA Prebiotic Matrix For Encapsulation of Probiotics: Physicochemical and Microbiological StudyAdriana CastellanosNo ratings yet
- DNA Barcodes On Their Own Are Not Enough To Describe A SpeciesDocument6 pagesDNA Barcodes On Their Own Are Not Enough To Describe A Speciesnscj.add.respNo ratings yet
- Nature 09201Document8 pagesNature 09201Felipe AlbrechtNo ratings yet
- 2008micron Fatbodiesofsemiengorged PDFDocument10 pages2008micron Fatbodiesofsemiengorged PDFAnonymous 75VkTFUtqNo ratings yet
- AgNPs Paper 2Document16 pagesAgNPs Paper 2VILEOLAGOLDNo ratings yet
- Lactobacillus SonuSinghAhirwaretal2017Document8 pagesLactobacillus SonuSinghAhirwaretal2017maira.myra19No ratings yet
- Diversity of Oral Micro Ora in Children of Age Group 6-12years in Pedodontics Department of RCDSRDocument7 pagesDiversity of Oral Micro Ora in Children of Age Group 6-12years in Pedodontics Department of RCDSRDiniNo ratings yet
- Article: Circulating Avian Influenza Viruses Closely Related To The 1918 Virus Have Pandemic PotentialDocument14 pagesArticle: Circulating Avian Influenza Viruses Closely Related To The 1918 Virus Have Pandemic PotentialAn TaraNo ratings yet
- Cholinesterase - Dibucaine: Quantitative Determination of Cholinesterase With Dibucaine InhibiciónDocument2 pagesCholinesterase - Dibucaine: Quantitative Determination of Cholinesterase With Dibucaine InhibiciónAldair VillegasNo ratings yet
- 10 1 1 925 8209 PDFDocument112 pages10 1 1 925 8209 PDFjpsingh75No ratings yet
- Ico Kin Etic S& Ico Dyn Am Ics: Professor Dr. Najat A. HasanDocument34 pagesIco Kin Etic S& Ico Dyn Am Ics: Professor Dr. Najat A. HasanMelkamu AyeNo ratings yet
- CSEC - Form 4 Human and Social Biology NotesDocument4 pagesCSEC - Form 4 Human and Social Biology NotesLeon Abel80% (5)
- Biology Sem 1 Exam PapersqDocument13 pagesBiology Sem 1 Exam Papersqpc usageNo ratings yet
- MCQS Microbiology For SendupDocument10 pagesMCQS Microbiology For SendupNab MughalNo ratings yet
- Scientific Background of Brahmu Muhurta PDFDocument3 pagesScientific Background of Brahmu Muhurta PDFLingareddyNo ratings yet
- BaSS 2019 Final ProgramDocument37 pagesBaSS 2019 Final ProgramArtanNo ratings yet
- 18btc101j Biochem-Unit 1Document134 pages18btc101j Biochem-Unit 1nikteshgNo ratings yet
- Tissue TypesDocument16 pagesTissue TypesJoanna FrancoNo ratings yet
- Photosynthesis and Cellular Respiration LabDocument2 pagesPhotosynthesis and Cellular Respiration Labapi-285366980No ratings yet
- 1physics-An Introduction SSC PresentationDocument28 pages1physics-An Introduction SSC PresentationPrincess ZoyaNo ratings yet
- BCH 332 Lectures 1 and 2 0Document33 pagesBCH 332 Lectures 1 and 2 0asdNo ratings yet
- Materi 5-Bioleaching Mineral OksidaDocument44 pagesMateri 5-Bioleaching Mineral OksidaVicky Faras Barunson PanggabeanNo ratings yet
- Hodgkin Lymphoma 1Document17 pagesHodgkin Lymphoma 1taufik perdanaNo ratings yet
- Solving Cold Cases With DNA Boston Strangler CaseDocument3 pagesSolving Cold Cases With DNA Boston Strangler CasePranjal SinghalNo ratings yet
- L RhamnosusDocument16 pagesL RhamnosusGonzalo Báez VNo ratings yet
- UntitledDocument27 pagesUntitledapi-53247190No ratings yet
- Detoxifiere Mercur Etc-GhidDocument175 pagesDetoxifiere Mercur Etc-GhidRadulescu EugenNo ratings yet
- Character Sheet - Modern - Call of Cthulhu V6 PDFDocument2 pagesCharacter Sheet - Modern - Call of Cthulhu V6 PDFEd LocardNo ratings yet
- CellDocument4 pagesCellPrincess MillanNo ratings yet
- Pattern Recognition - PresentationDocument25 pagesPattern Recognition - PresentationnurfarizaxNo ratings yet
- PracticeDocument9 pagesPracticeReuben John AbalayanNo ratings yet
- Processing of Temporomandibular Joint Dislocation Cases at Igd Rsud UNDATA PALU IN 2017-2018Document4 pagesProcessing of Temporomandibular Joint Dislocation Cases at Igd Rsud UNDATA PALU IN 2017-2018Jamalul HikmahNo ratings yet
- 1117 Microbiological Best Laboratory PracticesDocument11 pages1117 Microbiological Best Laboratory PracticesNgân TrịnhNo ratings yet
- Presentation On Ich Topics & Guidelines With A Special Reference ToDocument79 pagesPresentation On Ich Topics & Guidelines With A Special Reference ToVidyaNo ratings yet
- PRAKRIT™ - Sutra To ScienceDocument8 pagesPRAKRIT™ - Sutra To ScienceSita ShankarNo ratings yet
- Fate MapDocument5 pagesFate MapRitesh Singh Kharwar100% (1)
- Bioceramic Sealer ReviewDocument11 pagesBioceramic Sealer ReviewArdelia RaissaNo ratings yet