You might also like
- 2008 DNA MANS JDocument17 pages2008 DNA MANS JProf.Dr.SoHayla AttallaNo ratings yet
- Research Notes Random Amplified Polymorphic DNA Fingerprints For Identification of Species in Poultry Pa TeDocument3 pagesResearch Notes Random Amplified Polymorphic DNA Fingerprints For Identification of Species in Poultry Pa TeAnand DangreNo ratings yet
- The Personal Identification of Many Samples Recovered From Under The SeaDocument7 pagesThe Personal Identification of Many Samples Recovered From Under The SeajessicaNo ratings yet
- Detection of Meat Species Using Taqman Real-Time PCR AssaysDocument8 pagesDetection of Meat Species Using Taqman Real-Time PCR Assaysytngllc_197006458No ratings yet
- Differentiation of Cattle Species in Beef by PCR-RFLP of Mitochondrial and Satellite DNADocument5 pagesDifferentiation of Cattle Species in Beef by PCR-RFLP of Mitochondrial and Satellite DNAMet RizalNo ratings yet
- Analysis of Htra Gene From Zebrafish (Danio Rerio) : Murwantoko, Chio Oka and Masashi KawaichiDocument10 pagesAnalysis of Htra Gene From Zebrafish (Danio Rerio) : Murwantoko, Chio Oka and Masashi KawaichiHadiNYNo ratings yet
- 7554 13381 1 PB PDFDocument10 pages7554 13381 1 PB PDFRamla YaniNo ratings yet
- DNA Barcoding of Chinese Native Chicken Breeds Through COI GeneDocument8 pagesDNA Barcoding of Chinese Native Chicken Breeds Through COI Genemnipat007No ratings yet
- Repeat Regions in MT DnaDocument9 pagesRepeat Regions in MT DnaVeerendra UppalapatiNo ratings yet
- Submitted by Palash Ghosh M.SC Biotechnology Amity University, Haryana 2017Document29 pagesSubmitted by Palash Ghosh M.SC Biotechnology Amity University, Haryana 2017Palash GhoshNo ratings yet
- Fingerprinting Genomes Using PCR With Arbitrary PrimersDocument6 pagesFingerprinting Genomes Using PCR With Arbitrary PrimersraunakmsNo ratings yet
- Odriozola - Development and Validationof I-DNA1 - A 15-Loci Multiplex System For Identity TestingDocument10 pagesOdriozola - Development and Validationof I-DNA1 - A 15-Loci Multiplex System For Identity TestingspanishvcuNo ratings yet
- Forensic Animal DNA Analysis Using Economical Two-StepDocument10 pagesForensic Animal DNA Analysis Using Economical Two-StepFatima 999No ratings yet
- DiZinno Et Al., 2002 - Mitochondrial DNA Sequencing of Beetle Larvae (Omosita) Recovered From Human BoneDocument3 pagesDiZinno Et Al., 2002 - Mitochondrial DNA Sequencing of Beetle Larvae (Omosita) Recovered From Human BoneJean-Bernard HuchetNo ratings yet
- Artificial ChromosDocument12 pagesArtificial ChromosNada MohsenNo ratings yet
- 1723 UploadDocument5 pages1723 UploadbodhidharNo ratings yet
- Expressed Sequence Tag (EST) Analysis of A Schistosoma Japonicum Cercariae CDNA LibraryDocument10 pagesExpressed Sequence Tag (EST) Analysis of A Schistosoma Japonicum Cercariae CDNA LibraryahmicphdNo ratings yet
- 3287c PDFDocument7 pages3287c PDFChikondi TemboNo ratings yet
- Genetic AssessmentDocument22 pagesGenetic AssessmentW Montague Cobb LabNo ratings yet
- Science - Abo7257 SMDocument105 pagesScience - Abo7257 SM张议No ratings yet
- Edward 1991,1992 DNA Typing and Genetic Mapping With Trimeric and Tetrameric Tandem Repeats.Document11 pagesEdward 1991,1992 DNA Typing and Genetic Mapping With Trimeric and Tetrameric Tandem Repeats.Htein Lynn AungNo ratings yet
- Localization of Single Copy Gene by PRINS TechniqueDocument2 pagesLocalization of Single Copy Gene by PRINS TechniqueEAPNo ratings yet
- Forensic Science International 132 (2003) 76-81 PDFDocument6 pagesForensic Science International 132 (2003) 76-81 PDFJe RivasNo ratings yet
- 2017 - Sumer, N., Yldirimhan, H.S - DNA Sequencing of Digenea Nuclear lsrDNA of The Whiskered Brown Bat, Myotis Aurescens (Vespertilionidae Chiroptera), From Turkey PDFDocument4 pages2017 - Sumer, N., Yldirimhan, H.S - DNA Sequencing of Digenea Nuclear lsrDNA of The Whiskered Brown Bat, Myotis Aurescens (Vespertilionidae Chiroptera), From Turkey PDFStoian GoranovNo ratings yet
- Sequence Variation in Humans and Other Primates at Six Short Tandem Repeat Loci Used in Forensic Identity TestingDocument10 pagesSequence Variation in Humans and Other Primates at Six Short Tandem Repeat Loci Used in Forensic Identity TestingJose Manriquez NaveasNo ratings yet
- Cochlear Function in Mice With Only One Copy of The Prestin GeneDocument13 pagesCochlear Function in Mice With Only One Copy of The Prestin GeneAlex WongNo ratings yet
- Amer. J. Pathol.-2002-Urist-1199-1206Document8 pagesAmer. J. Pathol.-2002-Urist-1199-1206Charles J. DiComo, PhDNo ratings yet
- A Noninvasive, Direct Real Time PCR Method For Sex Determination in Multiple Avian SpeciesDocument3 pagesA Noninvasive, Direct Real Time PCR Method For Sex Determination in Multiple Avian SpeciesFatima 999No ratings yet
- DNA Typing From Skeletal Remains: Evaluation of Multiplex and Megaplex STR Systems On DNA Isolated From Bone and Teeth SamplesDocument6 pagesDNA Typing From Skeletal Remains: Evaluation of Multiplex and Megaplex STR Systems On DNA Isolated From Bone and Teeth SamplesFatima ColonelNo ratings yet
- Transcriptomic Analysis of Testicular Gene Expression in Normal and Cryptorchid HorsesDocument13 pagesTranscriptomic Analysis of Testicular Gene Expression in Normal and Cryptorchid Horsesalejandro sotoNo ratings yet
- A Comparative Analysis of Microscopy and PCR Based Detection Methods ForDocument6 pagesA Comparative Analysis of Microscopy and PCR Based Detection Methods ForBiswa Ranjan MaharanaNo ratings yet
- ITGβ6 and its association with FMD infection in crossbred cattle of KeralaDocument4 pagesITGβ6 and its association with FMD infection in crossbred cattle of KeralaAKHILA MRNo ratings yet
- Tesis PHD Rodríguez-HidalgoDocument139 pagesTesis PHD Rodríguez-HidalgoNiñoTorresNo ratings yet
- 2462.full TecbucaDocument2 pages2462.full TecbucaMauro Porcel de PeraltaNo ratings yet
- Flow CytometryDocument10 pagesFlow CytometryDaniel BarrigaNo ratings yet
- Cloning, Characterization, and Distribution of An mRNA Encoding A H + - ATPase Subunit in The Mantle of Pearl Oyster, Pinctada FucataDocument8 pagesCloning, Characterization, and Distribution of An mRNA Encoding A H + - ATPase Subunit in The Mantle of Pearl Oyster, Pinctada FucataGunawan MuhammadNo ratings yet
- 10 0000@cancerres Aacrjournals Org@content@60@6@1677Document7 pages10 0000@cancerres Aacrjournals Org@content@60@6@1677Hariadi Adha FNo ratings yet
- Identification of Scrapie Infection From Blood Serum by Fourier Transform Infrared SpectrosDocument4 pagesIdentification of Scrapie Infection From Blood Serum by Fourier Transform Infrared SpectrosKarina CordovaNo ratings yet
- Meat Science: S. Kunhareang, H. Zhou, J.G.H. HickfordDocument2 pagesMeat Science: S. Kunhareang, H. Zhou, J.G.H. HickfordPutu MahendraNo ratings yet
- Short Tandem RepeatsDocument15 pagesShort Tandem RepeatsJessica LeyvaNo ratings yet
- Sistem Otot Dan Kerangka UnggasDocument20 pagesSistem Otot Dan Kerangka UnggasWahyu Solhan MaghfurNo ratings yet
- Cucumber SSRe28Document5 pagesCucumber SSRe28Thuong HoNo ratings yet
- Simoncini T Naure 2000Document4 pagesSimoncini T Naure 2000DetaiACNo ratings yet
- A New Strategy For The Discrimination of Mitochondrial DNA Haplogroups in Han Population Yang JFS 2011Document5 pagesA New Strategy For The Discrimination of Mitochondrial DNA Haplogroups in Han Population Yang JFS 2011spanishvcuNo ratings yet
- Aw1435@txstate - Edu: Amanda Wilson #594648 Bio 2450 Genetics Dr. N. Martin Amanda SchultzDocument12 pagesAw1435@txstate - Edu: Amanda Wilson #594648 Bio 2450 Genetics Dr. N. Martin Amanda Schultzaw1435No ratings yet
- Significance of The Patched Gene in Developmental Biology: Evan Zhou T05 CMMB 403Document9 pagesSignificance of The Patched Gene in Developmental Biology: Evan Zhou T05 CMMB 403EvanNo ratings yet
- Genome Res.-2007-He-1471-7Document8 pagesGenome Res.-2007-He-1471-7Richard Icaro EsparzaNo ratings yet
- RBMO - Extraccion ADNDocument6 pagesRBMO - Extraccion ADNBlanca Idalia Gonzalez GarzaNo ratings yet
- Isolation and Expression of Two Distinct Sox8 Genes in Mudloach (Misgurnus Anguillicaudatus)Document16 pagesIsolation and Expression of Two Distinct Sox8 Genes in Mudloach (Misgurnus Anguillicaudatus)josekinNo ratings yet
- Arctigenin From Fructus Arctii Is A Novel Suppressor of Heat Shock Response in Mammalian CellsDocument8 pagesArctigenin From Fructus Arctii Is A Novel Suppressor of Heat Shock Response in Mammalian CellsDomitian PascaNo ratings yet
- Parade LaDocument6 pagesParade Laptrocha21No ratings yet
- Allele Frequencies of Six MiniSTR Loci in The Population of Northern PortugalDocument3 pagesAllele Frequencies of Six MiniSTR Loci in The Population of Northern PortugalspanishvcuNo ratings yet
- 2023 12 02 569703v1 FullDocument5 pages2023 12 02 569703v1 Fullcbrs.vetmedNo ratings yet
- PCR Detection of Tritrichomonas Foetus in Preputial Bull Uid Without Prior DNA IsolationDocument6 pagesPCR Detection of Tritrichomonas Foetus in Preputial Bull Uid Without Prior DNA IsolationYoga Mahendra PandiaNo ratings yet
- Detection of Toxoplasma Gondii by PCR and Quantitative PCR With High Specificty and Lower Limit of DetectionDocument7 pagesDetection of Toxoplasma Gondii by PCR and Quantitative PCR With High Specificty and Lower Limit of DetectionSholatya FurqonieNo ratings yet
- Wa0014.Document3 pagesWa0014.G.N. BhuvaneshwaranNo ratings yet
- 3897 LeptospiraDocument5 pages3897 LeptospiraFernanda Cordova HernandezNo ratings yet
- Myostatin Molecular Analysis of Ovine Myostatin GeDocument3 pagesMyostatin Molecular Analysis of Ovine Myostatin GeLe Minh Thanh 010086No ratings yet
- Evolutionary Parameters of The Transcribed Mammalian GenomeDocument6 pagesEvolutionary Parameters of The Transcribed Mammalian GenomeLau Mandy KMNo ratings yet
- Fast Facts: Comprehensive Genomic Profiling: Making precision medicine possibleFrom EverandFast Facts: Comprehensive Genomic Profiling: Making precision medicine possibleNo ratings yet
- Screen 2014 Uricchio 119 27Document9 pagesScreen 2014 Uricchio 119 27NazishTazeemNo ratings yet
- The Trend Trader Nick Radge On Demand PDFDocument8 pagesThe Trend Trader Nick Radge On Demand PDFDedi Tri LaksonoNo ratings yet
- Generation Transmission and Distribution Notes.Document88 pagesGeneration Transmission and Distribution Notes.Abhishek NirmalkarNo ratings yet
- The Light BulbDocument6 pagesThe Light Bulbapi-244765407No ratings yet
- Proceso Gmaw (RMD)Document8 pagesProceso Gmaw (RMD)Wilfredo RamirezNo ratings yet
- The Eminence in Shadow, Vol. 4Document255 pagesThe Eminence in Shadow, Vol. 4Ezra Salvame83% (6)
- Test Bank For Organizations Behavior Structure Processes 14th Edition James GibsonDocument36 pagesTest Bank For Organizations Behavior Structure Processes 14th Edition James Gibsonfurrygrindlett3t1bt100% (37)
- Research Propsal v1.2Document3 pagesResearch Propsal v1.2john7904No ratings yet
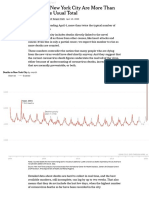
- Deaths in New York City Are More Than Double The Usual TotalDocument3 pagesDeaths in New York City Are More Than Double The Usual TotalRamón RuizNo ratings yet
- Structural Analysis of English SyntaxDocument26 pagesStructural Analysis of English SyntaxremovableNo ratings yet
- Asme B30.26 (2004)Document38 pagesAsme B30.26 (2004)Omar RamirezNo ratings yet
- BC Caribou Recovery Program Stakeholders Teleconference MinutesDocument5 pagesBC Caribou Recovery Program Stakeholders Teleconference MinutesRevelstoke MountaineerNo ratings yet
- Tikkun Kisay HaShemDocument47 pagesTikkun Kisay HaShemYochananMauritzHummasti100% (1)
- Hermes - GodDocument19 pagesHermes - GodAvocadoraNo ratings yet
- Aeroacoustic Optimization of Wind Turbine Airfoils by Combining Thermographic and Acoustic Measurement DataDocument4 pagesAeroacoustic Optimization of Wind Turbine Airfoils by Combining Thermographic and Acoustic Measurement DatamoussaouiNo ratings yet
- Too Early! by Anton Pavlovich ChekhovDocument4 pagesToo Early! by Anton Pavlovich Chekhovapi-19787590No ratings yet
- Tutorial 2 EtherchannelDocument3 pagesTutorial 2 EtherchannelOng CHNo ratings yet
- Make - Volume 13 - Volume 13Document212 pagesMake - Volume 13 - Volume 13Pedernal Lisiao100% (2)
- Transfer Request 2022 - CBOA CHENNAIDocument5 pagesTransfer Request 2022 - CBOA CHENNAIVignesh NarayananNo ratings yet
- Renaissance RapierDocument66 pagesRenaissance Rapierrshpr100% (1)
- Machine Design Key 2014Document15 pagesMachine Design Key 2014SouvikDasNo ratings yet
- Activity 1.1.2 Simple Machine Practice Problems KeyDocument6 pagesActivity 1.1.2 Simple Machine Practice Problems KeyNehemiah GriffinNo ratings yet
- Segment Reporting Decentralized Operations and Responsibility Accounting SystemDocument34 pagesSegment Reporting Decentralized Operations and Responsibility Accounting SystemalliahnahNo ratings yet
- Tea Board of India PDFDocument18 pagesTea Board of India PDFDebasish RazNo ratings yet
- Assignment 1 To 7Document23 pagesAssignment 1 To 7KashishNo ratings yet
- 6 Best Chainsaw Under 300: A Complete Buying Guide: What We LikeDocument5 pages6 Best Chainsaw Under 300: A Complete Buying Guide: What We LikeFahim WatsonNo ratings yet
- Uber+PM+Prioritisation+Assignment+Submission+File-Shashank KaranthDocument6 pagesUber+PM+Prioritisation+Assignment+Submission+File-Shashank KaranthShashank KaranthNo ratings yet
- Water Cement RatioDocument5 pagesWater Cement RatioCastro FarfansNo ratings yet
- English Grammar Class 2 Sentences and Phrases - Learn and Practice - Download Free PDFDocument8 pagesEnglish Grammar Class 2 Sentences and Phrases - Learn and Practice - Download Free PDFSlesha SinhaNo ratings yet
- Artificial Intelligence in Power System OperationsDocument8 pagesArtificial Intelligence in Power System OperationsDummyofindiaIndiaNo ratings yet