You might also like
- Aedes Species Ecological and Host Feeding Patterns in The North-Eastern Parts of South Africa, 2014-2018Document14 pagesAedes Species Ecological and Host Feeding Patterns in The North-Eastern Parts of South Africa, 2014-2018jimmyNo ratings yet
- Prevalence and Pattern of Waterborne Parasitic Infections in Eastern Africa A Systematic Scoping ReviewDocument8 pagesPrevalence and Pattern of Waterborne Parasitic Infections in Eastern Africa A Systematic Scoping ReviewWiwien HendrawanNo ratings yet
- Viruses 15 00636Document19 pagesViruses 15 00636skyggetrinnNo ratings yet
- Molecular Ecology - 2020 - Soghigian - Genetic Evidence For The Origin of Aedes Aegypti The Yellow Fever Mosquito in TheDocument14 pagesMolecular Ecology - 2020 - Soghigian - Genetic Evidence For The Origin of Aedes Aegypti The Yellow Fever Mosquito in TheGabriella StefanieNo ratings yet
- s12936 023 04815 9Document12 pagess12936 023 04815 9Hosanna AGBANGLANo ratings yet
- BIR 2019 Buxton EtalDocument7 pagesBIR 2019 Buxton EtalskyggetrinnNo ratings yet
- Epidemiology of Taenia Saginata TaeniosiscysticercDocument12 pagesEpidemiology of Taenia Saginata Taeniosiscysticercحسام الدين إسماعيلNo ratings yet
- Surveillance of Aedes Albopictus Skuse Breeding Preference in Selected Dengue Outbreak Localities, Peninsular MalaysiaDocument17 pagesSurveillance of Aedes Albopictus Skuse Breeding Preference in Selected Dengue Outbreak Localities, Peninsular MalaysiaAlflavius JNo ratings yet
- pntd.0005941 BOMDocument16 pagespntd.0005941 BOMhenriquesbimbeNo ratings yet
- Vector Incrimination and Transmission of Avian Malaria at An Aquarium in Japan: Mismatch in Parasite Composition Between Mosquitoes and PenguinsDocument12 pagesVector Incrimination and Transmission of Avian Malaria at An Aquarium in Japan: Mismatch in Parasite Composition Between Mosquitoes and Penguinsyowan wandikboNo ratings yet
- Anopheles MicrobiologyDocument4 pagesAnopheles MicrobiologyNatalyaNo ratings yet
- Identification of Wolbachia New Strains From Aedes Aegypti Mosquitoes, The Vector of Dengue Fever in Jeddah ProvinceDocument9 pagesIdentification of Wolbachia New Strains From Aedes Aegypti Mosquitoes, The Vector of Dengue Fever in Jeddah ProvinceFebby AuliaNo ratings yet
- Malacological Survey and Geographical Distribution of Vector Snails For Schistosomiasis Within Informal Settlements of Kisumu City, Western KenyaDocument9 pagesMalacological Survey and Geographical Distribution of Vector Snails For Schistosomiasis Within Informal Settlements of Kisumu City, Western KenyaJalison RêgoNo ratings yet
- The Incidence and Mortality of Yellow Fever in Africa: A Systematic Review and Meta-AnalysisDocument11 pagesThe Incidence and Mortality of Yellow Fever in Africa: A Systematic Review and Meta-AnalysisdanielavinuezaobandoNo ratings yet
- Malaria in North Africa: A Review of The Status of Vectors and ParasitesDocument13 pagesMalaria in North Africa: A Review of The Status of Vectors and ParasitesMichael JonesNo ratings yet
- Musca Domestica Acts As Transport Vector HostsDocument5 pagesMusca Domestica Acts As Transport Vector HostsIqbal ArdiansyahNo ratings yet
- ARTICLE Tiba1262023IJPR107398Document10 pagesARTICLE Tiba1262023IJPR107398tibaNo ratings yet
- CJ AlhassanDocument98 pagesCJ AlhassanDr K.O. ACHEMANo ratings yet
- UntitledDocument18 pagesUntitledFred ChibwanaNo ratings yet
- Knowledge, Awareness, Practice and Preventive Method Against Mosquito Borne Diseases in Tezpur, Assam, Northeast IndiaDocument4 pagesKnowledge, Awareness, Practice and Preventive Method Against Mosquito Borne Diseases in Tezpur, Assam, Northeast IndiaRabindra HazarikaNo ratings yet
- Anopheline Species Composition and The 1Document10 pagesAnopheline Species Composition and The 1IvanNo ratings yet
- Mapping The Burden of Cholera in Sub-Saharan Africa and Implications For Control: An Analysis of Data Across Geographical ScalesDocument8 pagesMapping The Burden of Cholera in Sub-Saharan Africa and Implications For Control: An Analysis of Data Across Geographical ScalesMasithaNo ratings yet
- Temporal and Spatial Distribution Characteristics in The Natural Plague Foci of Chinese Mongolian Gerbils Based On Spatial AutocorrelationDocument10 pagesTemporal and Spatial Distribution Characteristics in The Natural Plague Foci of Chinese Mongolian Gerbils Based On Spatial Autocorrelation121417skmNo ratings yet
- 1Document13 pages1yowan wandikboNo ratings yet
- NKOMBODocument13 pagesNKOMBOSadock KwizeraNo ratings yet
- Astuti2019 Article PaediatricDengueInfectionInCirDocument12 pagesAstuti2019 Article PaediatricDengueInfectionInCirfadhilNo ratings yet
- PandV Hardy-Geology DiscussionDocument16 pagesPandV Hardy-Geology DiscussionRachelNo ratings yet
- A Participatory Approach To Address Within Country Cross Border Malaria: The Case of Menoreh Hills in Java, IndonesiaDocument9 pagesA Participatory Approach To Address Within Country Cross Border Malaria: The Case of Menoreh Hills in Java, Indonesiayowan wandikboNo ratings yet
- International Journal of Infectious Diseases: EditorialDocument5 pagesInternational Journal of Infectious Diseases: Editorialblooom_00No ratings yet
- Dónde Se Presentará La Nueva PandemiaDocument8 pagesDónde Se Presentará La Nueva PandemiajuanvillicanaNo ratings yet
- Galley Proof - 2023 - IJPR - 107398Document10 pagesGalley Proof - 2023 - IJPR - 107398tibaNo ratings yet
- Plant Height and Lifespan Predict Range Size in Southern African GrassesDocument14 pagesPlant Height and Lifespan Predict Range Size in Southern African GrassesAriadne Cristina De AntonioNo ratings yet
- Ebola Virus Disease: A Manual for EVD ManagementFrom EverandEbola Virus Disease: A Manual for EVD ManagementMarta LadoNo ratings yet
- Urbanization Impact On Mosquito Community and The Transmission Potential of Filarial Infection in Central EuropeDocument10 pagesUrbanization Impact On Mosquito Community and The Transmission Potential of Filarial Infection in Central EuropeGustavo AndresNo ratings yet
- Widespread Occurrence of The Amphibian CDocument8 pagesWidespread Occurrence of The Amphibian Cabeje kassieNo ratings yet
- GeographicallocationDocument6 pagesGeographicallocationسوسو توتوNo ratings yet
- Antenatal CareDocument6 pagesAntenatal Carenanadewi892No ratings yet
- Prevalence and Intensity of Urinary Schistosomiasis in Weweso M/A Basic School-Kumasi MetropolisDocument60 pagesPrevalence and Intensity of Urinary Schistosomiasis in Weweso M/A Basic School-Kumasi MetropolisDennis Isaac AmoahNo ratings yet
- Schistosomiasis in ChildrenDocument8 pagesSchistosomiasis in ChildrenarghamgedeNo ratings yet
- Asian Citrus Psyllid: Biology, Ecology and Management of the Huanglongbing VectorFrom EverandAsian Citrus Psyllid: Biology, Ecology and Management of the Huanglongbing VectorJawwad A QureshiNo ratings yet
- 230941-Article Text-560492-1-10-20220831Document19 pages230941-Article Text-560492-1-10-20220831fireworkNo ratings yet
- GovoetchanDocument12 pagesGovoetchanibrahima1968No ratings yet
- Acta Tropica: SciencedirectDocument7 pagesActa Tropica: SciencedirectDr-Sadaqat Ali RaoNo ratings yet
- Knowledge, Compliance and Effects of The Oral Cholera Vaccine at The Hill Top of The Ndobo Community, Littoral Region, Douala, CameroonDocument6 pagesKnowledge, Compliance and Effects of The Oral Cholera Vaccine at The Hill Top of The Ndobo Community, Littoral Region, Douala, CameroonInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- A Study To Evaluvate The Effectiveness of Structured Teaching Programme On Knowledge Regarding Prevention of Masquito Borne Diseases Among Rural People of MuchakhandiDocument5 pagesA Study To Evaluvate The Effectiveness of Structured Teaching Programme On Knowledge Regarding Prevention of Masquito Borne Diseases Among Rural People of MuchakhandiInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- The Global Distribution and Transmission Limits of Lymphatic Filariasis: Past and PresentDocument20 pagesThe Global Distribution and Transmission Limits of Lymphatic Filariasis: Past and PresentNanda AzzahraNo ratings yet
- s12917 023 03766 2Document20 pagess12917 023 03766 2yudhi arjentiniaNo ratings yet
- Biosafety and Health: SciencedirectDocument7 pagesBiosafety and Health: SciencedirectIslam Ausraf RajibNo ratings yet
- Geospatial Modeled Analysis and Laboratory Based Technology For Determination of Malaria Risk and Burden in A Rural CommunityDocument14 pagesGeospatial Modeled Analysis and Laboratory Based Technology For Determination of Malaria Risk and Burden in A Rural CommunitySegunNo ratings yet
- Drones Against Vector-Borne Diseases: FocusDocument3 pagesDrones Against Vector-Borne Diseases: Focusyowan wandikboNo ratings yet
- Phytoprotection: Almouner A.A. Yattara, Amadou K. Coulibaly and Frédéric FrancisDocument8 pagesPhytoprotection: Almouner A.A. Yattara, Amadou K. Coulibaly and Frédéric Francismouhamed sowNo ratings yet
- Low Practice of Malaria Prevention Among Migrants and Seasonal Farmworkers in Metema and West Armacheho Districts, Northwest EthiopiaDocument9 pagesLow Practice of Malaria Prevention Among Migrants and Seasonal Farmworkers in Metema and West Armacheho Districts, Northwest EthiopiaDaniel BoibanNo ratings yet
- Pathogens 10 00190Document16 pagesPathogens 10 00190Mariana BarcelóNo ratings yet
- Detection of Novel Coronaviruses in Bats in Myanmar: A1111111111 A1111111111 A1111111111 A1111111111 A1111111111Document11 pagesDetection of Novel Coronaviruses in Bats in Myanmar: A1111111111 A1111111111 A1111111111 A1111111111 A1111111111VoskiNo ratings yet
- Document PDFDocument13 pagesDocument PDFPororoNo ratings yet
- Global Prevalence of ASCARIS LUMBRICOIDESDocument16 pagesGlobal Prevalence of ASCARIS LUMBRICOIDESprofnutrialecuriNo ratings yet
- Socioeconomic and Human Behavioral Factors Associated With Aedes Aegypti (Diptera: Culicidae) Immature Habitat in Tucson, AZDocument9 pagesSocioeconomic and Human Behavioral Factors Associated With Aedes Aegypti (Diptera: Culicidae) Immature Habitat in Tucson, AZFaiz ZulkifliNo ratings yet
- Malaria and ChildrenDocument11 pagesMalaria and ChildrenCARLES MSOLANo ratings yet
- Global Risk Mapping For Major Ae.Document11 pagesGlobal Risk Mapping For Major Ae.Jorge VillalobosNo ratings yet
- The Dominant Anopheles Vectors of Human Malaria in Africa, Europe and The Middle East: Occurrence Data, Distribution Maps and Bionomic PrécisDocument34 pagesThe Dominant Anopheles Vectors of Human Malaria in Africa, Europe and The Middle East: Occurrence Data, Distribution Maps and Bionomic Précisibrahima1968No ratings yet
- Microbial Biomass ProductionDocument38 pagesMicrobial Biomass ProductionSanaullahSunnyNo ratings yet
- INICET JULY 2021 Solved Paper PDF PathologyDocument36 pagesINICET JULY 2021 Solved Paper PDF Pathologysusan291No ratings yet
- ch20-22 - Probleme SetDocument8 pagesch20-22 - Probleme SetReese VespertineNo ratings yet
- Serotonin Reuptake InhibitorsDocument6 pagesSerotonin Reuptake Inhibitorsdsekulic_1No ratings yet
- Evolution by Polyploidy: Dan GraurDocument36 pagesEvolution by Polyploidy: Dan GraurSelvaraju ParthibhanNo ratings yet
- BIOL 3162 Lab 2-Introduction To Microbial Biotech Techniques ADocument7 pagesBIOL 3162 Lab 2-Introduction To Microbial Biotech Techniques Arube10000No ratings yet
- Recombinant DNA Technology LectureDocument50 pagesRecombinant DNA Technology LectureJean Carmelette BalalloNo ratings yet
- Elijah James - 35-DNA Replication Worksheet - CompleteDocument4 pagesElijah James - 35-DNA Replication Worksheet - CompleteElijah JamesNo ratings yet
- SNASTI-ICCS, 14-18.: Daftar PustakaDocument4 pagesSNASTI-ICCS, 14-18.: Daftar PustakaPradhani Fakhira DhaneswariNo ratings yet
- COVID-19 Pathology Accepted PrePrint PDFDocument25 pagesCOVID-19 Pathology Accepted PrePrint PDFkambangNo ratings yet
- Study Guide - Behavioral Ecology: Short AnswerDocument7 pagesStudy Guide - Behavioral Ecology: Short Answerowls_1102No ratings yet
- 6253 Johnson QuiviraDocument101 pages6253 Johnson QuiviraLalo Morales Muñoz100% (2)
- GENBIO2Document17 pagesGENBIO2Perlyn Del Pilar OduyaNo ratings yet
- Prevention of HerpatitisDocument54 pagesPrevention of Herpatitisapi-270822363No ratings yet
- Course Foundation Biology I Code BIO091 Lecturer / Tutor Duration 1 HOUR 30 Minutes Student Name Nur Syafiqah Najihah BT Abdul Aziz ID GroupDocument9 pagesCourse Foundation Biology I Code BIO091 Lecturer / Tutor Duration 1 HOUR 30 Minutes Student Name Nur Syafiqah Najihah BT Abdul Aziz ID GroupNur Syafiqah NajihahNo ratings yet
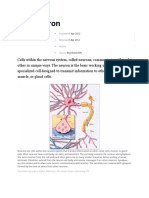
- The Neuron: Published Reviewed Author SourceDocument10 pagesThe Neuron: Published Reviewed Author SourceJennie KimNo ratings yet
- Biological Sequence Databases: A. National Center For Biotechnology Information (NCBI)Document41 pagesBiological Sequence Databases: A. National Center For Biotechnology Information (NCBI)Isha ChopraNo ratings yet
- Origin of LifeDocument4 pagesOrigin of LifeBatak CyberTeamNo ratings yet
- The Cell Cycle and Cancer 2011Document28 pagesThe Cell Cycle and Cancer 2011Rajesh BalakrishnanNo ratings yet
- Lesson-09 - BACTERIAL CULTURE Media PDFDocument8 pagesLesson-09 - BACTERIAL CULTURE Media PDFawadsonNo ratings yet
- Experiment Phototropism and GeotropismDocument3 pagesExperiment Phototropism and GeotropismPramudith Liyanage100% (3)
- PapadogiannisDocument10 pagesPapadogiannisjhupressNo ratings yet
- 1195VET - Topic 5 - Types of Gene ActionDocument13 pages1195VET - Topic 5 - Types of Gene Actionاحمد البزورNo ratings yet
- 9671 41590 1 PBDocument4 pages9671 41590 1 PBNicolas De la cruz CasadiegoNo ratings yet
- Harrsion 1991 PDFDocument8 pagesHarrsion 1991 PDFrozh rasulNo ratings yet
- History of MicrobiologyDocument3 pagesHistory of MicrobiologyAngeli ButedNo ratings yet
- Clin. Microbiol. Rev.-1999-Guarro-fungal Taxonomy 1999Document47 pagesClin. Microbiol. Rev.-1999-Guarro-fungal Taxonomy 1999cesiahdezNo ratings yet
- GST Fusion BrochureDocument12 pagesGST Fusion BrochureDolphingNo ratings yet
- Acs Jcim 2c01400Document8 pagesAcs Jcim 2c01400谢吴辰No ratings yet
- Biotech CosmeticsDocument3 pagesBiotech CosmeticsAnonymous Ptxr6wl9DhNo ratings yet