You might also like
- The Energy Codes: Dr. Sue MorterDocument19 pagesThe Energy Codes: Dr. Sue MorterVeres Beatrix92% (12)
- Answer Diagnosis: 1. RhythmDocument2 pagesAnswer Diagnosis: 1. RhythmSuggula Vamsi KrishnaNo ratings yet
- Case Study Presentation FroehlichDocument27 pagesCase Study Presentation Froehlichapi-499098722100% (1)
- Adult Dehydration Guideline Final 2015Document3 pagesAdult Dehydration Guideline Final 2015Brillyant S RaintamaNo ratings yet
- Local Nursing Licensure ExamDocument14 pagesLocal Nursing Licensure Examnikko042790No ratings yet
- Review: Genomic Approaches To Studying The Human MicrobiotaDocument7 pagesReview: Genomic Approaches To Studying The Human MicrobiotaJing XueNo ratings yet
- The Bactericidal Action of Penicillin: New Clues To An Unsolved MysteryDocument5 pagesThe Bactericidal Action of Penicillin: New Clues To An Unsolved MysteryJhordy Alexis Flores SantiagoNo ratings yet
- Cole Et Al 2021. The Surface Lipoproteins of Gram-Negative Bacteria - Protectors and Foragers in Harsh EnvironmentsDocument15 pagesCole Et Al 2021. The Surface Lipoproteins of Gram-Negative Bacteria - Protectors and Foragers in Harsh EnvironmentsAlmendra MosqueraNo ratings yet
- Club 04Document13 pagesClub 04Muñoz Cerón Smiht YhoissNo ratings yet
- Chronic Bacterial Infections: Living With Unwanted Guests: Douglas Young, Tracy Hussell and Gordon DouganDocument7 pagesChronic Bacterial Infections: Living With Unwanted Guests: Douglas Young, Tracy Hussell and Gordon DouganMarcus ViníciusNo ratings yet
- Toxins of Enteric BacteriaDocument5 pagesToxins of Enteric BacteriaKangan KhannaNo ratings yet
- Role of Microbiota in Immunity and InflammationDocument12 pagesRole of Microbiota in Immunity and InflammationGerald John PazNo ratings yet
- Role of The Gut Microbiota in Immunity and Inflammatory DiseaseDocument15 pagesRole of The Gut Microbiota in Immunity and Inflammatory DiseaseNatália LopesNo ratings yet
- Coba Browser Baru Dengan Terjemahan Otomatis.: Plaque WhenDocument5 pagesCoba Browser Baru Dengan Terjemahan Otomatis.: Plaque WhenDiana SuhartiNo ratings yet
- Infeccion MicrobianaDocument18 pagesInfeccion MicrobianaRosemary Neto AlfonsoNo ratings yet
- Host Microbiomes and Their Interactions With Immune SystemDocument9 pagesHost Microbiomes and Their Interactions With Immune SystemGerald John PazNo ratings yet
- Kamada2013 PDFDocument15 pagesKamada2013 PDFÂngela Souza BarroqueiroNo ratings yet
- The Microbiome and Regulation of Mucosal ImmunityDocument8 pagesThe Microbiome and Regulation of Mucosal ImmunityLevente BalázsNo ratings yet
- Host-Pathogen Interactions: The Seduction of Molecular Cross TalkDocument7 pagesHost-Pathogen Interactions: The Seduction of Molecular Cross TalkPoppy Siska IsabellaNo ratings yet
- tmp3159 TMPDocument7 pagestmp3159 TMPFrontiersNo ratings yet
- Evasion InmuneDocument8 pagesEvasion Inmunenydiacastillom2268No ratings yet
- Bacterial Virulence Factors: Secreted For Survival: Mycobacterium TuberculosisDocument10 pagesBacterial Virulence Factors: Secreted For Survival: Mycobacterium TuberculosisRahayuNo ratings yet
- Rodriguez-Vega 2021Document15 pagesRodriguez-Vega 2021Hugo RodriguesNo ratings yet
- Inmune 19Document9 pagesInmune 19Pablo Pinaya SotomayorNo ratings yet
- The Microbiome and Innate ImmunityDocument10 pagesThe Microbiome and Innate ImmunityGabriel HernándezNo ratings yet
- Bacteriilogy Lecture Notes (2) : Bacteria: Structure-Function-Pathogenicity RelationshipsDocument19 pagesBacteriilogy Lecture Notes (2) : Bacteria: Structure-Function-Pathogenicity RelationshipsNa KhanNo ratings yet
- A Gonzalez18originDocument26 pagesA Gonzalez18originRara IndiraNo ratings yet
- Jurnal Sains Dan Kesehatan: Peranan Sel Limfosit Dalam Imunulogi: Artikel ReviewDocument13 pagesJurnal Sains Dan Kesehatan: Peranan Sel Limfosit Dalam Imunulogi: Artikel ReviewDearma KhairunnisaNo ratings yet
- Lactobacillus Fermentum ME-3 - An Antimicrobial and Antioxidative ProbioticDocument27 pagesLactobacillus Fermentum ME-3 - An Antimicrobial and Antioxidative ProbioticMemyselfNo ratings yet
- Chapter 2 and 3 ImmunologyDocument16 pagesChapter 2 and 3 ImmunologyRevathyNo ratings yet
- Identification, Subtyping and Virulence Determination of Listeria Monocytogenes, An Important Foodborne PathogenDocument15 pagesIdentification, Subtyping and Virulence Determination of Listeria Monocytogenes, An Important Foodborne PathogenDiuThiDoNo ratings yet
- Communications in Oral Biofilm: IjcrrDocument5 pagesCommunications in Oral Biofilm: IjcrrLutfia NirwanaNo ratings yet
- The Basis For The Immunoregulatory Role of Macrophages and Other Accessory Cells.Document7 pagesThe Basis For The Immunoregulatory Role of Macrophages and Other Accessory Cells.Cristian Gutiérrez VeraNo ratings yet
- Molecular Mechanisms Used by Salmonella To Evade The Immune SystemDocument35 pagesMolecular Mechanisms Used by Salmonella To Evade The Immune SystemANDREA FERNANDA MUÑOZ NARVAEZNo ratings yet
- Artis 2008Document10 pagesArtis 2008contramondum1232039No ratings yet
- 1 s2.0 S2173580817300445 MainDocument10 pages1 s2.0 S2173580817300445 MainEvangelina RivarolaNo ratings yet
- Lecture 1 - Immunology Online Questions Chapter 1Document15 pagesLecture 1 - Immunology Online Questions Chapter 1DrBhawna AroraNo ratings yet
- Host Immune Response To Chlamydia InfectionDocument17 pagesHost Immune Response To Chlamydia Infectionapi-266835840No ratings yet
- CytokinesDocument12 pagesCytokinesclventuriniNo ratings yet
- Salmonellae Interplay With Host Cells: Andrea Haraga, Maikke B. Ohlson and Samuel I. MillerDocument14 pagesSalmonellae Interplay With Host Cells: Andrea Haraga, Maikke B. Ohlson and Samuel I. MillerShaznay Dela CruzNo ratings yet
- Abraham and Medzhitov 2011 - Interactions Between The Host Innate Immune System and Microbes in Inflammatory Bowel DiseaseDocument9 pagesAbraham and Medzhitov 2011 - Interactions Between The Host Innate Immune System and Microbes in Inflammatory Bowel DiseaseAchille BroggiNo ratings yet
- Entamoeba Histolytica: Review ArticleDocument11 pagesEntamoeba Histolytica: Review ArticleDian IslamiatiNo ratings yet
- Review: Role of The Microbiota in Immunity and InflammationDocument21 pagesReview: Role of The Microbiota in Immunity and InflammationJULIANA VITORIANo ratings yet
- Immunity Lec 3Document11 pagesImmunity Lec 3محمد حسين عبد الهاديNo ratings yet
- Breaking Down BiofilmsDocument3 pagesBreaking Down Biofilmsleila.hiloutNo ratings yet
- Does The Buck Stop With The Bugs?: An Overview of Microbial Dysbiosis in Rheumatoid ArthritisDocument13 pagesDoes The Buck Stop With The Bugs?: An Overview of Microbial Dysbiosis in Rheumatoid ArthritiswiwekaNo ratings yet
- Cei0161 0378 PDFDocument11 pagesCei0161 0378 PDFErik Jaya GunawanNo ratings yet
- Intestinal Microbiota and Sclerosis Lateral AmyotrophicDocument11 pagesIntestinal Microbiota and Sclerosis Lateral AmyotrophicaluminiossaolucasNo ratings yet
- Harshada and PranavDocument13 pagesHarshada and PranavPranav NakhateNo ratings yet
- He Pa To BiliaryDocument14 pagesHe Pa To BiliaryMayerly Camila González CórdobaNo ratings yet
- The Microbial WorldDocument32 pagesThe Microbial WorldNurul Khairiyah IINo ratings yet
- Projeto de IC - Inflamação e LeishmaniaDocument1 pageProjeto de IC - Inflamação e LeishmaniaDiovana RamosNo ratings yet
- Salmonellae Interactions With Host Process NATUREDocument15 pagesSalmonellae Interactions With Host Process NATUREDiegoNo ratings yet
- Intestinal Epithelial Cells Regulators of Barrier Function and Immune HomeostasisDocument13 pagesIntestinal Epithelial Cells Regulators of Barrier Function and Immune HomeostasisCristian Gutiérrez VeraNo ratings yet
- Panniculitis: Adipose Tissue and ImmunityDocument7 pagesPanniculitis: Adipose Tissue and ImmunityAnnisa Chaerani BurhanuddinNo ratings yet
- Gut: Key Element On Immune System Regulation: Review - Human and Animal HealthDocument14 pagesGut: Key Element On Immune System Regulation: Review - Human and Animal HealthyutefupNo ratings yet
- Connecting The Immune System, Systemic Chronic Inflammation and The Gut Microbiome - The Role of Sex.Document23 pagesConnecting The Immune System, Systemic Chronic Inflammation and The Gut Microbiome - The Role of Sex.MariaAndreaLaraSalasNo ratings yet
- Contamination of Tissue Cultures by Mycoplasmas: Shlomo Rottem, Nechama S. Kosower and Jonathan D. KornspanDocument24 pagesContamination of Tissue Cultures by Mycoplasmas: Shlomo Rottem, Nechama S. Kosower and Jonathan D. KornspanNarasimha MurthyNo ratings yet
- J. Biol. Chem.-2019-Coelho-1202-17 PDFDocument17 pagesJ. Biol. Chem.-2019-Coelho-1202-17 PDFJoel Torres VillenaNo ratings yet
- Enteric Bacteria Boost Defences Against Oxidative Stress in Entamoeba HistolyticaDocument12 pagesEnteric Bacteria Boost Defences Against Oxidative Stress in Entamoeba Histolyticagwyneth.green.512No ratings yet
- Male and Female InfertilityDocument164 pagesMale and Female InfertilityNkopengieh Rene PochiekenNo ratings yet
- Virulence Factors of Candida SpeciesDocument6 pagesVirulence Factors of Candida Specieslaura_ruiz_99No ratings yet
- Microbial Virulence Factors: Molecular SciencesDocument6 pagesMicrobial Virulence Factors: Molecular SciencesAnasNo ratings yet
- Microbiota Overview and Implication in IDocument15 pagesMicrobiota Overview and Implication in IJULIETANo ratings yet
- The Role of Products of the Histocompatibility Gene Complex in Immune ResponsesFrom EverandThe Role of Products of the Histocompatibility Gene Complex in Immune ResponsesNo ratings yet
- Gida Profiling Tool PDFDocument4 pagesGida Profiling Tool PDFRaym AvilaNo ratings yet
- Universidad Peruana Cayetano Heredia: Facultad de Medicina Veterinaria y ZootecniaDocument48 pagesUniversidad Peruana Cayetano Heredia: Facultad de Medicina Veterinaria y ZootecniaJose Luis Jaen RamosNo ratings yet
- Diarrhea PDFDocument14 pagesDiarrhea PDFabegail eliahNo ratings yet
- Área C Ingeniería: Simulacro Virtual Acem Semestral - Repaso San Marcos 2021-IDocument26 pagesÁrea C Ingeniería: Simulacro Virtual Acem Semestral - Repaso San Marcos 2021-IPERCY VITON DIAZNo ratings yet
- Medication Deferral ListDocument2 pagesMedication Deferral ListKwiila KaoNo ratings yet
- Insulin and Oral Hypoglycemic DrugsDocument45 pagesInsulin and Oral Hypoglycemic DrugsRamadi PrameelaNo ratings yet
- Termossonicação Nondairy ResultadosDocument9 pagesTermossonicação Nondairy ResultadosBruna VieiraNo ratings yet
- Tuberculosis DrugsDocument136 pagesTuberculosis DrugsSyed Gulshan NaqviNo ratings yet
- The Dural Tail Sign in The Diagnosis of MeningiomasDocument6 pagesThe Dural Tail Sign in The Diagnosis of MeningiomasCabinet VeterinarNo ratings yet
- A. Loss of Deep Tendon ReflexesDocument15 pagesA. Loss of Deep Tendon Reflexes2013SecBNo ratings yet
- JKGT - usakti,+Journal+manager,+Manuscript+JKGT-21-07 18 R1Document4 pagesJKGT - usakti,+Journal+manager,+Manuscript+JKGT-21-07 18 R1felitaNo ratings yet
- Caring For Pts W Dementia - SBDocument25 pagesCaring For Pts W Dementia - SBriley liuNo ratings yet
- Vereckei TVDocument12 pagesVereckei TVVallejo Romo Alberto CristianNo ratings yet
- Concept Essay IdeasDocument10 pagesConcept Essay Ideasd3gpmvqw100% (2)
- Hiv Informative Genre FinalDocument9 pagesHiv Informative Genre Finalapi-510346047No ratings yet
- HIV - AIDS PresentationDocument17 pagesHIV - AIDS PresentationphilNo ratings yet
- Brain TumorDocument1 pageBrain TumorChase XerolfNo ratings yet
- Drug Study CCMHDocument35 pagesDrug Study CCMHJose Mari F. EsguerraNo ratings yet
- Enhanced Recovery After Intestinal Surgery (ERAS)Document4 pagesEnhanced Recovery After Intestinal Surgery (ERAS)Andre F SusantioNo ratings yet
- Respiratory System Pathology (Part 1)Document8 pagesRespiratory System Pathology (Part 1)chrisp7No ratings yet
- CPC 0 Series QuestionsDocument6 pagesCPC 0 Series Questions19ucs175No ratings yet
- Improving Mental Healthcare in the Re-Entry System ActDocument18 pagesImproving Mental Healthcare in the Re-Entry System ActerickieferNo ratings yet
- Create PDFDocument88 pagesCreate PDFSisnawaty SabatinyNo ratings yet
- Food and Chemical Toxicology: Can Ali Agca, Mehmet Tuzcu, Armagan Hayirli, Kazim SahinDocument6 pagesFood and Chemical Toxicology: Can Ali Agca, Mehmet Tuzcu, Armagan Hayirli, Kazim SahinΔημητρης ΦιλιογλουNo ratings yet
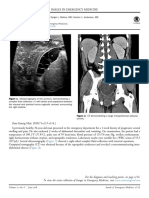
- 0196-0644/$-See Front Matter: 71, No. 6: June 2018 Annals of Emergency MedicineDocument2 pages0196-0644/$-See Front Matter: 71, No. 6: June 2018 Annals of Emergency MedicineTeja Laksana NukanaNo ratings yet