You might also like
- Fgene 11 00546Document14 pagesFgene 11 00546Karla MontesNo ratings yet
- Heliyon Hemp Article 2022 (1327)Document11 pagesHeliyon Hemp Article 2022 (1327)S R Shahriar SazidNo ratings yet
- Suman Rawte and Ritu R. SaxenaDocument8 pagesSuman Rawte and Ritu R. SaxenaCleimerNo ratings yet
- A Taxonomic Study of Twelve Wild Forage Species of Fabaceae - 2021 - HeliyonDocument12 pagesA Taxonomic Study of Twelve Wild Forage Species of Fabaceae - 2021 - HeliyonTONY VILCHEZ YARIHUAMANNo ratings yet
- 1-s2.0-S025462991633650X-mainDocument9 pages1-s2.0-S025462991633650X-maininna50237No ratings yet
- 1 s2.0 S025462991633650X MainDocument9 pages1 s2.0 S025462991633650X MainAnny LovibNo ratings yet
- Rose GeneDocument15 pagesRose GeneBekele TilahunNo ratings yet
- Applications of molecular markers in genetic diversity studies of maizeDocument8 pagesApplications of molecular markers in genetic diversity studies of maizeBlaxez YTNo ratings yet
- Role of Mol Markers in SWDocument10 pagesRole of Mol Markers in SWAlexia MihalceaNo ratings yet
- Exploring The Molecular Basis of Heterosis For Plant BreedingDocument12 pagesExploring The Molecular Basis of Heterosis For Plant BreedingMaría Del Carmen Sanches IvancichNo ratings yet
- MARCADORES[1]Document9 pagesMARCADORES[1]Carlos Alberto Alva SifuentesNo ratings yet
- Assessment of Genetic Diversity Population StructuDocument26 pagesAssessment of Genetic Diversity Population StructuAnny LovibNo ratings yet
- 1.1.3 Genetic Diversity Analysis in Lathyrus Sativus Using SSR Molecular Markers 2Document8 pages1.1.3 Genetic Diversity Analysis in Lathyrus Sativus Using SSR Molecular Markers 2RITESH CHANDANo ratings yet
- chandrawati2014Document8 pageschandrawati2014inna50237No ratings yet
- 3RBBCoriandisArthropods20198117 31Document16 pages3RBBCoriandisArthropods20198117 31silverelemental24No ratings yet
- DNA Fingerprinting and Phylogenetic Relationship of The Genus Chlorophytum Ker Gawl, From India Using AFLPDocument7 pagesDNA Fingerprinting and Phylogenetic Relationship of The Genus Chlorophytum Ker Gawl, From India Using AFLPEditor IJTSRDNo ratings yet
- 1 s2.0 S0304423820305318 MainDocument12 pages1 s2.0 S0304423820305318 Mainnasrul hakimNo ratings yet
- Dila 1Document5 pagesDila 1NirwanaNo ratings yet
- SABRAO J Breed Genet 49 2 192 200 Rini 1Document10 pagesSABRAO J Breed Genet 49 2 192 200 Rini 1MANSYUR ANDRAWINSYAH HSNo ratings yet
- Assessment of Genetic Diversity in Crop Plants - An OverviewDocument8 pagesAssessment of Genetic Diversity in Crop Plants - An OverviewEyasuNo ratings yet
- Kang Et Al 2014. Adaptative and Nonadaptative Genome SizeDocument11 pagesKang Et Al 2014. Adaptative and Nonadaptative Genome SizelaurachalupNo ratings yet
- DUS Charactetrization Article MSCDocument13 pagesDUS Charactetrization Article MSCK.amarnathNo ratings yet
- PDFDocument13 pagesPDFAricatul JannahNo ratings yet
- Changes in Soil Bacteria Functional Ecology Associated With Morchella Rufobrunnea Fruiting in A Natural HabitatDocument12 pagesChanges in Soil Bacteria Functional Ecology Associated With Morchella Rufobrunnea Fruiting in A Natural HabitatEzra M. OrlofskyNo ratings yet
- horticulturae-10-00120 (1)Document16 pageshorticulturae-10-00120 (1)omaragronomieNo ratings yet
- Genome Sizes in Diploid and Allopolyploid Arachis L. SpeciesDocument18 pagesGenome Sizes in Diploid and Allopolyploid Arachis L. SpeciesSofi VazquezNo ratings yet
- Genetic Divergence in Soybean (Glycine Max L. Merill) During Rabi SeasonDocument3 pagesGenetic Divergence in Soybean (Glycine Max L. Merill) During Rabi SeasonMandalapu PallaviNo ratings yet
- 726 7391 1 PBDocument9 pages726 7391 1 PBpavithraNo ratings yet
- Molecular Diversity of Egyptian Cotton Gossypium BDocument8 pagesMolecular Diversity of Egyptian Cotton Gossypium BBenson TrentNo ratings yet
- IJHST Volume9 Issue1 Pages97-116Document21 pagesIJHST Volume9 Issue1 Pages97-116Novi anitaNo ratings yet
- Variability and Heritability Studies in Winged Bean (Psophocarpus Tetragonolobus (L.) DC.) For Seed Yield and Component Characters.Document4 pagesVariability and Heritability Studies in Winged Bean (Psophocarpus Tetragonolobus (L.) DC.) For Seed Yield and Component Characters.Journal of Environment and Bio-SciencesNo ratings yet
- (23279834 - HortScience) Amplified Fragment Length Polymorphism Analysis Provides Strategies For Improvement of Bitter Gourd (Momordica Charantia L.)Document7 pages(23279834 - HortScience) Amplified Fragment Length Polymorphism Analysis Provides Strategies For Improvement of Bitter Gourd (Momordica Charantia L.)Ralph OndrasNo ratings yet
- s10722-021-01202-8Document22 pagess10722-021-01202-8inna50237No ratings yet
- Ijasft 5 143Document6 pagesIjasft 5 143peertechzNo ratings yet
- 1 s2.0 S2214540015000298 MainDocument8 pages1 s2.0 S2214540015000298 MainmuhamadNo ratings yet
- 10 1111@boj 12442Document14 pages10 1111@boj 12442johnheverthNo ratings yet
- Estimating the Genetic Diversity of Ethiopian Noug (Guizotia Abyssinica (L.f.) Genotypes Using SSR MarkerDocument8 pagesEstimating the Genetic Diversity of Ethiopian Noug (Guizotia Abyssinica (L.f.) Genotypes Using SSR MarkerbirhanuNo ratings yet
- Biodiversidad GeneticaDocument10 pagesBiodiversidad GeneticaLuis Alberto Reyna CulquiNo ratings yet
- 23712-Article Text-71162-1-10-20211130Document9 pages23712-Article Text-71162-1-10-20211130nurmiyati unsNo ratings yet
- Genetic Analysisof Important Qualitativein RiceDocument6 pagesGenetic Analysisof Important Qualitativein RiceDeepak RajNo ratings yet
- CAJPSI - Volume 1 - Issue 4 - Pages 192-200 PDFDocument9 pagesCAJPSI - Volume 1 - Issue 4 - Pages 192-200 PDFKERTÉSZMÉRNÖK SZIEMKKNo ratings yet
- Genetic Diversity of A Maize Association Population With Restricted PhenologyDocument12 pagesGenetic Diversity of A Maize Association Population With Restricted PhenologyFrontiersNo ratings yet
- PAPER6_rosado2018Document10 pagesPAPER6_rosado2018diegoppxxNo ratings yet
- An Overview of "Insect Biodiversity"Document5 pagesAn Overview of "Insect Biodiversity"OscarNo ratings yet
- 2193 1801 2 313Document6 pages2193 1801 2 313Kartikeya TiwariNo ratings yet
- Diversity and Distributions - 2022 - Arranz - Genogeographic Clustering To Identify Cross Species Concordance of SpatialDocument13 pagesDiversity and Distributions - 2022 - Arranz - Genogeographic Clustering To Identify Cross Species Concordance of SpatialLetmestudyNo ratings yet
- Coccinia - 2012Document7 pagesCoccinia - 2012Suhara BeevyNo ratings yet
- Joprv22aug10 NusaibahDocument8 pagesJoprv22aug10 NusaibahbenzzzxNo ratings yet
- VariolaDocument11 pagesVariolaDAYANNE PEREIRA DE SOUZANo ratings yet
- Evolution of Floral Morphology and Symmetry in The Miconieae (Melastomataceae) : Multiple Generalization Trends Within A Specialized FamilyDocument16 pagesEvolution of Floral Morphology and Symmetry in The Miconieae (Melastomataceae) : Multiple Generalization Trends Within A Specialized FamilyDanielaTorresGarciaNo ratings yet
- Genetic Analysis and Character Association in Different - Solanki - 2015Document11 pagesGenetic Analysis and Character Association in Different - Solanki - 2015NurmalitaNo ratings yet
- +2018 Vol50no3rad12 GENETIKA MORFOLOSKEDocument12 pages+2018 Vol50no3rad12 GENETIKA MORFOLOSKEIrena CalicNo ratings yet
- GJESM Volume 7 Issue 3 Pages 473-484Document12 pagesGJESM Volume 7 Issue 3 Pages 473-484GJESMNo ratings yet
- 50 56PhyuPhyuDocument8 pages50 56PhyuPhyuIPPALA NARENDRA REDDY 2003721No ratings yet
- Morphological and Molecular Studies Reveal Phylogenetic Relationships Among Three Boerhavia SpeciesDocument13 pagesMorphological and Molecular Studies Reveal Phylogenetic Relationships Among Three Boerhavia SpeciesGustavo Andres Fuentes RodriguezNo ratings yet
- ManseguiaopaperDocument7 pagesManseguiaopaperTris MeNo ratings yet
- Barley Asian Journal of Biotechnology and Genetic EngeneeringDocument9 pagesBarley Asian Journal of Biotechnology and Genetic EngeneeringKadri AlineNo ratings yet
- Elife 68623 v2Document28 pagesElife 68623 v2MarNo ratings yet
- Identification of Variants Induced by Physical and Chemical Mutagens in Tuberose (Polianthes Tuberosa L.)Document6 pagesIdentification of Variants Induced by Physical and Chemical Mutagens in Tuberose (Polianthes Tuberosa L.)mohapatra1996No ratings yet
- Chapter 3 PowerPointDocument73 pagesChapter 3 PowerPointJohn NikolaevichNo ratings yet
- Lecture-13: Chapter - 3: Introduction of PharmacodynamicsDocument3 pagesLecture-13: Chapter - 3: Introduction of Pharmacodynamicsবাংলা দেশNo ratings yet
- Cahier D'anglais - TerminaleDocument4 pagesCahier D'anglais - TerminaleKanade SatoNo ratings yet
- General Biology Exam ReviewerDocument31 pagesGeneral Biology Exam ReviewerRayyyNo ratings yet
- 7.016 Recitation 17 - Fall 2018: Summary of Lectures 25 (11/9) & 26 (11/14)Document9 pages7.016 Recitation 17 - Fall 2018: Summary of Lectures 25 (11/9) & 26 (11/14)Shubham singhNo ratings yet
- RP96 44Document31 pagesRP96 44Serdar Abdulkerim GulliNo ratings yet
- Biochemistry Laboratory Modern Theory and Techniques-2nd EdDocument78 pagesBiochemistry Laboratory Modern Theory and Techniques-2nd EdCharles EdwardNo ratings yet
- JCJ BiosocialSLIntegration ResearchGateDocument21 pagesJCJ BiosocialSLIntegration ResearchGatesundesh kumarNo ratings yet
- Can Genes Predict Athletic Performance?Document2 pagesCan Genes Predict Athletic Performance?Channy PhamNo ratings yet
- On Lesson 14 Nucleic Acids and LipidsDocument40 pagesOn Lesson 14 Nucleic Acids and LipidsKamto EzenwamaduNo ratings yet
- Whole-Genome Shotgun Sequencing - Overview, Steps and AchievementsDocument8 pagesWhole-Genome Shotgun Sequencing - Overview, Steps and Achievementshunk gargNo ratings yet
- Global Developmental Delay GDDDocument2 pagesGlobal Developmental Delay GDDYanuar Adi SanjayaNo ratings yet
- Metabolic Fingerprints of Human Primary Endothelial and Fibroblast CellsDocument12 pagesMetabolic Fingerprints of Human Primary Endothelial and Fibroblast CellsDavid Galiano LatorreNo ratings yet
- 7.015 Intro Biology Course Covers Microbiology, ImmunologyDocument16 pages7.015 Intro Biology Course Covers Microbiology, ImmunologyLong NguyenNo ratings yet
- Introduction To EnymesDocument34 pagesIntroduction To EnymesMeshal NoorNo ratings yet
- Estimation of Genetic Variance On Tea (Camellia Sinensis (L.) O. Kuntze) Clones Based On Morphological Markers in Southwestern EthiopiaDocument7 pagesEstimation of Genetic Variance On Tea (Camellia Sinensis (L.) O. Kuntze) Clones Based On Morphological Markers in Southwestern EthiopiaPremier PublishersNo ratings yet
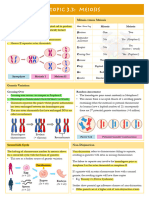
- 3.3 MeiosisDocument1 page3.3 MeiosisMaeva SeneNo ratings yet
- WhysexDocument4 pagesWhysexapi-31866583862% (13)
- CADD Drug Design StrategiesDocument22 pagesCADD Drug Design StrategiesMaleeha HussainNo ratings yet
- Detecting Prey From DNA in Predator Scats A Comparison With Morphological Analysis, Using Arctocephalus Seals Fed A Known DietDocument11 pagesDetecting Prey From DNA in Predator Scats A Comparison With Morphological Analysis, Using Arctocephalus Seals Fed A Known DietVladimir Delgado RodriguezNo ratings yet
- CL 10 Bio PracticalDocument4 pagesCL 10 Bio PracticalRasode me Kaun thaNo ratings yet
- Natural History of Highly Pathogenic Avian in Uenza H5N1: Virus ResearchDocument16 pagesNatural History of Highly Pathogenic Avian in Uenza H5N1: Virus ResearchAn TaraNo ratings yet
- 2014 YALCIN BinnazDocument107 pages2014 YALCIN BinnazMaaz NasimNo ratings yet
- MicrobiologyDocument4 pagesMicrobiologyHannah Grace CorveraNo ratings yet
- B.SC III Microbiology 2020-21Document26 pagesB.SC III Microbiology 2020-21Shardul ManeNo ratings yet
- Effectiveness of Mugwort in Treating Stomach Ache 2Document15 pagesEffectiveness of Mugwort in Treating Stomach Ache 2Shiela LalagunaNo ratings yet
- QP May-2017 Paper 2 PDFDocument20 pagesQP May-2017 Paper 2 PDFMoazzem HossainNo ratings yet
- Potanc-2 1Document23 pagesPotanc-2 1paskug2No ratings yet
- Part 1: Muscle Physiology: Skeletal MusclesDocument7 pagesPart 1: Muscle Physiology: Skeletal MusclesMyrtle Yvonne RagubNo ratings yet
- The Organic Codes An Introduction To Semantic BiolDocument3 pagesThe Organic Codes An Introduction To Semantic BiolIsmailNo ratings yet
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- Why We Sleep: Unlocking the Power of Sleep and DreamsFrom EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsRating: 4.5 out of 5 stars4.5/5 (2083)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (4)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)










![MARCADORES[1]](https://imgv2-2-f.scribdassets.com/img/document/723443164/149x198/0ea7a49e31/1713237771?v=1)