You might also like
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5811)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- Biography of Sanatan Goswami PDFDocument31 pagesBiography of Sanatan Goswami PDFDavor VeseljkoNo ratings yet
- Et Oxilow 20 Feed v01Document2 pagesEt Oxilow 20 Feed v01manuelenecp100% (2)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (844)
- Solucionario Problemas de Generadores DCDocument11 pagesSolucionario Problemas de Generadores DCELISBAN CHILLITUPA SEGOVIA100% (1)
- Todas Dengue 2018 Narrative ReportDocument3 pagesTodas Dengue 2018 Narrative ReportBOBOT JOEL ROMERONo ratings yet
- Piura y Sus Danzas Más Representativas - OdtDocument3 pagesPiura y Sus Danzas Más Representativas - OdtMel UpsNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
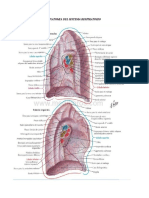
- Anatomia Del Sistema RespiratorioDocument7 pagesAnatomia Del Sistema RespiratorioJohn Feliciano CedeñoNo ratings yet
- Historia de Los Mundiales de FutbolDocument15 pagesHistoria de Los Mundiales de FutbolJohn Feliciano CedeñoNo ratings yet
- Examen Del RectoDocument20 pagesExamen Del RectoJohn Feliciano CedeñoNo ratings yet
- ANTIHIPERTENSIVOSDocument44 pagesANTIHIPERTENSIVOSJohn Feliciano CedeñoNo ratings yet
- Bradicardía SinusualDocument2 pagesBradicardía SinusualJohn Feliciano CedeñoNo ratings yet
- DIURETICOSDocument88 pagesDIURETICOSJohn Feliciano CedeñoNo ratings yet
- 21ST Century ReviewerDocument4 pages21ST Century ReviewerRhea Stephanie OliverosNo ratings yet
- French 3am19 3trim2Document2 pagesFrench 3am19 3trim2slimane100% (1)
- Forbes Estates LipaDocument19 pagesForbes Estates LipaNico SilvanoNo ratings yet
- Oncenio de Augusto LeguiaDocument3 pagesOncenio de Augusto LeguiaWalter Quiroz miravalNo ratings yet
- 19 Jan 24 Time Table Cse & CA Even Sem 2023-24Document161 pages19 Jan 24 Time Table Cse & CA Even Sem 2023-24officialpritam1504No ratings yet
- PREGUNTAS IntermedioDocument12 pagesPREGUNTAS IntermedioAntonio100% (1)
- Onboarding 2023Document68 pagesOnboarding 2023Cenk ErdincNo ratings yet
- Aula 1 Introdução À Pesquisa de MarketingDocument18 pagesAula 1 Introdução À Pesquisa de MarketingAdilson L. de BarrosNo ratings yet
- Remove BloatwareDocument7 pagesRemove BloatwareIpsrs ElektromedisNo ratings yet
- Separta HominimosDocument1 pageSeparta HominimosHenry RicaldiNo ratings yet
- Email To Manager Asking For Leave For Brother's Wedding (In The Middle of An Important Project)Document4 pagesEmail To Manager Asking For Leave For Brother's Wedding (In The Middle of An Important Project)Aryan NegiNo ratings yet
- Solucionario Evaluaciones Bilingüe 2ºDocument14 pagesSolucionario Evaluaciones Bilingüe 2ºniovekNo ratings yet
- Sharpe, J. (1993) - Historia Desde AbajoDocument3 pagesSharpe, J. (1993) - Historia Desde AbajoFelipilloDigitalNo ratings yet
- Danikas Christos Georgia 1971 GreeceDocument3 pagesDanikas Christos Georgia 1971 Greecethe missions networkNo ratings yet
- Topical Corticosteroid Use During Pregnancy: Motherisk UpdateDocument2 pagesTopical Corticosteroid Use During Pregnancy: Motherisk UpdateKhairul AlvaroNo ratings yet
- Guia INDECI FINAL Por Paginas PDFDocument100 pagesGuia INDECI FINAL Por Paginas PDFJavier Fernando Rodriguez RiegaNo ratings yet
- Sárgabarack FajtákDocument28 pagesSárgabarack Fajtákr4kemberNo ratings yet
- Trabajo de KickingballDocument24 pagesTrabajo de KickingballTonya Gates63% (16)
- Performance 1Document1 pagePerformance 1jarlrb4eNo ratings yet
- Ectopico Acog 2018Document10 pagesEctopico Acog 2018LAURA MELISSA GAVIRIA SEGURANo ratings yet
- Plan Estrategico para Mejora de Calidad de CafeDocument95 pagesPlan Estrategico para Mejora de Calidad de CafeerikaNo ratings yet
- Manual de Instruções - Labstar Zeta - 15202586 - AgrivalleDocument38 pagesManual de Instruções - Labstar Zeta - 15202586 - AgrivalleGabriel Vilas BoasNo ratings yet
- Instalación y Uso Básico de Aurman - Linux GNU BlogDocument3 pagesInstalación y Uso Básico de Aurman - Linux GNU BlogShashin KuroNo ratings yet
- Business Proposal For Using Fintech in Banking ArenaDocument4 pagesBusiness Proposal For Using Fintech in Banking ArenawoosanNo ratings yet
- Spray-Dried Honey PowderDocument1 pageSpray-Dried Honey PowderbiaNo ratings yet