You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Frequency in HZDocument63 pagesFrequency in HZPrashant DahaleNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- PSY 101 Sample Exam QuestionsDocument15 pagesPSY 101 Sample Exam Questionsveledimaraz80% (5)
- Neural Network PPT PresentationDocument23 pagesNeural Network PPT PresentationSiddharth Patel50% (2)
- Neurological Exam Checklist: Mental Status, Motor Skills, Sensory TestsDocument2 pagesNeurological Exam Checklist: Mental Status, Motor Skills, Sensory Testssara ashari100% (2)
- GUILLAIN BARRE SYNDROME CASE STUDY (Group)Document28 pagesGUILLAIN BARRE SYNDROME CASE STUDY (Group)Kylie Golindang100% (7)
- Engineering Unit Conversion PDFDocument5 pagesEngineering Unit Conversion PDFBarcepandoNo ratings yet
- Journal Role of Downstream SignallingDocument9 pagesJournal Role of Downstream Signallingherrbede4645No ratings yet
- ASIO4ALL v2 Instruction ManualDocument9 pagesASIO4ALL v2 Instruction Manualjosu100% (2)
- Marketing 1Document15 pagesMarketing 1herrbede4645No ratings yet
- Reflections (Remembering and Forgetting)Document1 pageReflections (Remembering and Forgetting)ShaniceNo ratings yet
- Vertigo Diagnosis and Management - 230425 - 212729Document56 pagesVertigo Diagnosis and Management - 230425 - 212729Annisa MuflikhasariNo ratings yet
- Prognosis Negative: Psychiatry and The Foibles of The Diagnostic and Statistical Manual V (DSM-V)Document7 pagesPrognosis Negative: Psychiatry and The Foibles of The Diagnostic and Statistical Manual V (DSM-V)Rekha KumariNo ratings yet
- NeuropeptidesDocument62 pagesNeuropeptidesshankarkjs1983No ratings yet
- Santiago Ramón y Cajal en La NeuroscienciaDocument11 pagesSantiago Ramón y Cajal en La NeuroscienciaAmy Medina VelazquezNo ratings yet
- Autism DSM IV Vs DSM-VDocument14 pagesAutism DSM IV Vs DSM-VNikita KarinNo ratings yet
- 15-Sensory OrgansDocument23 pages15-Sensory OrgansMilad HabibiNo ratings yet
- Banich Exec FunctionDocument6 pagesBanich Exec FunctionPaula PorrasNo ratings yet
- L Basic Neuroscience Lecture 13Document17 pagesL Basic Neuroscience Lecture 13monster40lbsNo ratings yet
- Chapter 7 - Our Attention Is Limited - Our Memory Is ImperfectDocument8 pagesChapter 7 - Our Attention Is Limited - Our Memory Is ImperfectSabin Manuela Kerwien LopezNo ratings yet
- Nursing Care Plan TB MeningitisDocument2 pagesNursing Care Plan TB Meningitisderic74% (19)
- Intraneuronal Signalling Pathways: Chair Person: Dr. S. Kiran Kumar Presenter: Dr. G - Sangha MitraDocument20 pagesIntraneuronal Signalling Pathways: Chair Person: Dr. S. Kiran Kumar Presenter: Dr. G - Sangha Mitrasangha_mitra_2No ratings yet
- Ortho History Taking & Physical ExaminationDocument28 pagesOrtho History Taking & Physical ExaminationCabdi WaliNo ratings yet
- Spinalbifida 121213120324 Phpapp02Document28 pagesSpinalbifida 121213120324 Phpapp02Silvana María Espinoza CuadrosNo ratings yet
- Limb Apraxias - The Influence of Higher Order Perceptual and Semantic Deficits in Motor Recovery After StrokeDocument14 pagesLimb Apraxias - The Influence of Higher Order Perceptual and Semantic Deficits in Motor Recovery After StrokeAdiel OjedaNo ratings yet
- NEURO Myasthenia GravisDocument25 pagesNEURO Myasthenia GravisNasrah N. MusaNo ratings yet
- Why Is An EEG Performed?Document4 pagesWhy Is An EEG Performed?Sai MuraliNo ratings yet
- Impaired Verbal CommunicationDocument3 pagesImpaired Verbal CommunicationChenee Mabulay100% (1)
- (Brief Discussion of The Lesson, If Possible Cite Examples) : SUBJECT (Uppercase and Bold)Document33 pages(Brief Discussion of The Lesson, If Possible Cite Examples) : SUBJECT (Uppercase and Bold)KD DelayunNo ratings yet
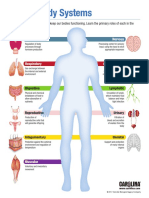
- Human Body System InfographicDocument12 pagesHuman Body System InfographicDiana VillanuevaNo ratings yet
- Juan Emilio Cuesta XII - Turing B Anatomy and Physiology Case Study #3Document5 pagesJuan Emilio Cuesta XII - Turing B Anatomy and Physiology Case Study #3Antonio NoblezaNo ratings yet
- Science Reviewer Q3 G10Document4 pagesScience Reviewer Q3 G10Jenny Ann SantosNo ratings yet
- Brain Gate Technology (Documentation)Document34 pagesBrain Gate Technology (Documentation)SrūJãn ChinnabathiniNo ratings yet
- Integumentary Sensory Organs in CrocodiliansDocument10 pagesIntegumentary Sensory Organs in CrocodiliansEduardo LeónNo ratings yet
- What Are The Differences Between Long-Term, Short-Term, and Working MemoryDocument15 pagesWhat Are The Differences Between Long-Term, Short-Term, and Working MemoryAsad KhanNo ratings yet