You might also like
- Leon Chaitow - Fascial Dysfunction 2ed PDFDocument407 pagesLeon Chaitow - Fascial Dysfunction 2ed PDFRukaphuong100% (12)
- Hema 1 August 2021Document84 pagesHema 1 August 2021Renz Gerard Amor100% (1)
- Pollination Class 12 Biology ProjectDocument21 pagesPollination Class 12 Biology ProjectASHOK SAGAR85% (20)
- Pollination Class 12 Biology ProjectDocument18 pagesPollination Class 12 Biology ProjectRajat Rawat0% (1)
- Physiology of PainDocument43 pagesPhysiology of PainNashwan ANo ratings yet
- Hormonal ImbalanceDocument4 pagesHormonal ImbalanceSeeraphNo ratings yet
- Pollination Class 12 Biology ProjectDocument18 pagesPollination Class 12 Biology ProjectSiddhartha Gautama76% (89)
- SWH AQA Revision Guide Biology 1Document42 pagesSWH AQA Revision Guide Biology 1afshinNo ratings yet
- Physiology of LarynxDocument115 pagesPhysiology of LarynxKashish HatukaNo ratings yet
- MSC Botany Paper-VIII Unit-8Document12 pagesMSC Botany Paper-VIII Unit-8løzanNo ratings yet
- Goats Reproduction Estrual Cycle Estrous Synchronization Artificial InseminationDocument11 pagesGoats Reproduction Estrual Cycle Estrous Synchronization Artificial InseminationBayu Anke MardianNo ratings yet
- Pollen-Pistil Interaction: Pollen Attachment and HydrationDocument6 pagesPollen-Pistil Interaction: Pollen Attachment and HydrationSreeja Raj100% (1)
- Unit 11Document33 pagesUnit 11shiva kumar goudNo ratings yet
- Vmi2011 757194Document13 pagesVmi2011 757194Ahmad SolihinNo ratings yet
- Bioengineering and The Ovarian FollicleDocument7 pagesBioengineering and The Ovarian FollicleViji ThulasiramanNo ratings yet
- Clarke 2017Document22 pagesClarke 2017Isabela LopesNo ratings yet
- CAP 2 Regulation of Follicle Formation andDocument27 pagesCAP 2 Regulation of Follicle Formation andgilson_pessoaNo ratings yet
- Reproductive Biology and EndocrinologyDocument7 pagesReproductive Biology and EndocrinologyFabiola Ramos CavalieriNo ratings yet
- 1 s2.0 S0093691X13003865 MainDocument7 pages1 s2.0 S0093691X13003865 Maincarrerajm22No ratings yet
- Topological Progression in Proliferating Epithelia Is Driven by A Unique Variation in Polygon DistributionDocument8 pagesTopological Progression in Proliferating Epithelia Is Driven by A Unique Variation in Polygon DistributionAuriNo ratings yet
- Plant Cell Environment - 2007 - VIOLA - Symplastic Connection Is Required For Bud Outgrowth Following Dormancy in PotatoDocument11 pagesPlant Cell Environment - 2007 - VIOLA - Symplastic Connection Is Required For Bud Outgrowth Following Dormancy in PotatoPaolaHanccoLandaNo ratings yet
- Andrade 2019Document12 pagesAndrade 2019Isabela LopesNo ratings yet
- KvmzupollinationprojectDocument18 pagesKvmzupollinationprojectK. K KumarNo ratings yet
- Meyer Et Al. 2004Document15 pagesMeyer Et Al. 2004Aline Freitas de MeloNo ratings yet
- Pollen Germination and Tube GrowthDocument40 pagesPollen Germination and Tube Growthabiandabicementwork arumugamNo ratings yet
- PollinationDocument22 pagesPollinationNAVEEN DHIMAN100% (1)
- Oocyte Control of Ovarian Follicular Development and Function in MammalsDocument10 pagesOocyte Control of Ovarian Follicular Development and Function in MammalsArDi EcHaNo ratings yet
- Reproductive Biology and Technology in Animals PDFDocument126 pagesReproductive Biology and Technology in Animals PDFJuan CaroNo ratings yet
- JournalDocument5 pagesJournalGabby RachediaNo ratings yet
- POLLINATIONDocument16 pagesPOLLINATIONAlejandro Jose RebuaNo ratings yet
- Regulation of Blastocyst Formation.Document24 pagesRegulation of Blastocyst Formation.christopher LopezNo ratings yet
- Hamedi 2017Document22 pagesHamedi 2017joaovictorandre0609No ratings yet
- Ashley - REPRODUCTIVE DEVELOPMENT OF THE FEMALEDocument17 pagesAshley - REPRODUCTIVE DEVELOPMENT OF THE FEMALEPEMUDA RABBANINo ratings yet
- Cortical Granule FunctionDocument17 pagesCortical Granule FunctionEric PhillipsNo ratings yet
- 2010 Vital Et AlDocument12 pages2010 Vital Et Alsbd2hugNo ratings yet
- Pollination WikipediaDocument12 pagesPollination WikipediaAnirban MallickNo ratings yet
- Oocyte Maturation: A Story of Arrest and Release: Frontiers in Bioscience (Scholar Edition) January 2013Document55 pagesOocyte Maturation: A Story of Arrest and Release: Frontiers in Bioscience (Scholar Edition) January 2013ArinilHaqueNo ratings yet
- Department of Biology: Jawahar Navodaya Vidyalaya ShegaonDocument25 pagesDepartment of Biology: Jawahar Navodaya Vidyalaya Shegaonprasad panjabrao mapariNo ratings yet
- Domestic Animal Endocrinology: F.R.C.L. Almeida, A.L.N. Alvarenga DiasDocument9 pagesDomestic Animal Endocrinology: F.R.C.L. Almeida, A.L.N. Alvarenga DiasPigostilNo ratings yet
- BlastulaDocument4 pagesBlastulaCyanDesNo ratings yet
- Pollination in Sunflower and LegumDocument18 pagesPollination in Sunflower and Legumkanha kumarNo ratings yet
- Zhao2019 - Invited Review Accelerating Mammary Gland InvolutionDocument17 pagesZhao2019 - Invited Review Accelerating Mammary Gland Involutionabdelaziz CHELIGHOUMNo ratings yet
- SPERMATOGENESIS The First Wave of Spermatogenesis Moreno 2018Document6 pagesSPERMATOGENESIS The First Wave of Spermatogenesis Moreno 2018LudimilaNo ratings yet
- 3684-Texto Del Manuscrito Completo (Cuadros y Figuras Insertos) - 21783-2-10-20181117Document9 pages3684-Texto Del Manuscrito Completo (Cuadros y Figuras Insertos) - 21783-2-10-20181117Dhamaris Sharai Gonzalez SaldivarNo ratings yet
- Implantation and PlacentationDocument45 pagesImplantation and PlacentationsoniyaarpNo ratings yet
- Biology Investigatory ProjectDocument19 pagesBiology Investigatory ProjectPriyanks Rout50% (2)
- Choice BoardDocument5 pagesChoice BoardHAIZA HANIM BINTI ABD AZIZ MoeNo ratings yet
- The Use of Zygotic Embryos As Explants For in Vitro Propagation: An OverviewDocument27 pagesThe Use of Zygotic Embryos As Explants For in Vitro Propagation: An OverviewCesarRamNo ratings yet
- The Mathematics of Sexual Attraction: MinireviewDocument5 pagesThe Mathematics of Sexual Attraction: MinireviewNuthal SrinivasanNo ratings yet
- Signalling Pathways Implicated in Early Mammary Gland Morphogenesis and Breast CancerDocument19 pagesSignalling Pathways Implicated in Early Mammary Gland Morphogenesis and Breast CancerhajimehikariNo ratings yet
- JCB 202102146Document15 pagesJCB 202102146Flavio NavarreteNo ratings yet
- Advances in Animal in Vitro FertilizationDocument33 pagesAdvances in Animal in Vitro FertilizationTaha TottiNo ratings yet
- SAQ EmbryologyDocument9 pagesSAQ EmbryologyShilpa DuttaNo ratings yet
- Pcos 3Document15 pagesPcos 3Deepak KumarNo ratings yet
- Club 04Document13 pagesClub 04Muñoz Cerón Smiht YhoissNo ratings yet
- The Prevention of Aging in MammalsDocument5 pagesThe Prevention of Aging in MammalsJeremy KnippleNo ratings yet
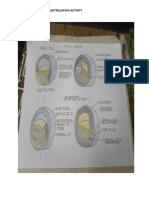
- GASTRULATION-ACTIVITY - DEV-BIO - Bio 102Document6 pagesGASTRULATION-ACTIVITY - DEV-BIO - Bio 102Quentin LopezNo ratings yet
- 6 FertilizationDocument30 pages6 FertilizationPuran BistaNo ratings yet
- Mucus ServixDocument10 pagesMucus ServixHaykal Estu BhismoroNo ratings yet
- Biology Project: A Cbse English Medium Co-Educational Senior Secondary School SESSION: 2021-2022Document10 pagesBiology Project: A Cbse English Medium Co-Educational Senior Secondary School SESSION: 2021-2022Omar Abdullah100% (1)
- Robert 2022Document15 pagesRobert 2022Isabela LopesNo ratings yet
- FERTILIZATIONDocument7 pagesFERTILIZATIONMarcos AlvarezNo ratings yet
- Module 5Document10 pagesModule 5Phan MhiveNo ratings yet
- Grimm 2020Document16 pagesGrimm 2020MD LarasatiNo ratings yet
- Biology 10 00238 v3Document13 pagesBiology 10 00238 v3michaelj5No ratings yet
- BIOMOLECULES-CHEMISTRY ProjectsDocument24 pagesBIOMOLECULES-CHEMISTRY ProjectsAjay KumarNo ratings yet
- Healing and RepairDocument38 pagesHealing and RepairDr.rajender ThakralNo ratings yet
- 2021 Anatomy Respiratory Module BCQsDocument12 pages2021 Anatomy Respiratory Module BCQsAmsha MangiNo ratings yet
- Characteristics & Classification of Living Organisms (Multiple Choice) 2 QPDocument8 pagesCharacteristics & Classification of Living Organisms (Multiple Choice) 2 QPAdetohunNo ratings yet
- Nursing Care For Patients With Digestive Disorder IntoxicationDocument8 pagesNursing Care For Patients With Digestive Disorder Intoxicationyanti yulianaNo ratings yet
- B1.1 Knowledge OrganiserDocument3 pagesB1.1 Knowledge OrganisermtauhidNo ratings yet
- Dubai Seafex 2013 Fuari TalepleriDocument10 pagesDubai Seafex 2013 Fuari Talepleritapioca leNo ratings yet
- Histology of Female Reproductive SystemDocument11 pagesHistology of Female Reproductive SystemYoshua AdhisaktiNo ratings yet
- Cap. 01 - Amphibian Anatomy and PhysiologyDocument12 pagesCap. 01 - Amphibian Anatomy and PhysiologyNailson JúniorNo ratings yet
- Odontoblasts: The Cells Forming and Maintaining DentineDocument7 pagesOdontoblasts: The Cells Forming and Maintaining DentineJoniMukaNo ratings yet
- Balanced Salt SolutionDocument36 pagesBalanced Salt Solutionnaili nsnNo ratings yet
- Cell Structure and FunctionDocument44 pagesCell Structure and FunctionPedro SuyuNo ratings yet
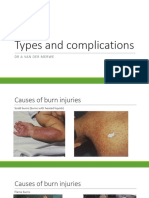
- Types and Complications Burn (Physiotherapy)Document26 pagesTypes and Complications Burn (Physiotherapy)JaniNo ratings yet
- Full Download Book Middletons Allergy Essentials 1E PDFDocument41 pagesFull Download Book Middletons Allergy Essentials 1E PDFbarry.schwend504100% (20)
- H3P2 - Nur Hanani - Lab Report SB025Document6 pagesH3P2 - Nur Hanani - Lab Report SB025Nur HananiNo ratings yet
- Cardiac Cycle: DR Rida Ajmal KhanDocument29 pagesCardiac Cycle: DR Rida Ajmal KhanMooma fatimaNo ratings yet
- TTPB7THESTORYOFAUXIN ShortDocument65 pagesTTPB7THESTORYOFAUXIN ShortDiksha GahlotNo ratings yet
- Home WDocument38 pagesHome WSriram SriramNo ratings yet
- Xylem: A Vascular Tissue in Land PlantsDocument4 pagesXylem: A Vascular Tissue in Land PlantsJoshua-Jonai JulyeNo ratings yet
- 1584special Sample Paper 2Document10 pages1584special Sample Paper 2Internal VibesNo ratings yet
- Human Factors ChadezDocument242 pagesHuman Factors ChadezChristopher MutyambiziNo ratings yet
- EI ShipraDocument7 pagesEI ShipraShipra BharadwajNo ratings yet
- Sensation and PerceptionDocument44 pagesSensation and PerceptionAarohi PatilNo ratings yet
- GENERAL ANAESTHESIA - Presentation Editted RecordedDocument31 pagesGENERAL ANAESTHESIA - Presentation Editted RecordedTatenda SibandaNo ratings yet