You might also like
- 41 Food and Thought DesktopDocument360 pages41 Food and Thought DesktopVera Whitehead100% (1)
- TTAD 3 Volume Transcripts CompleteDocument103 pagesTTAD 3 Volume Transcripts CompleteRaul Cortes100% (1)
- The Kidney Gut Axis Implications For Nutrition Care 2015 Megan RossiDocument5 pagesThe Kidney Gut Axis Implications For Nutrition Care 2015 Megan RossiWobik HopeNo ratings yet
- Understanding Normal and Clinical Nutrition 11th Edition by Sharon Rady Rolfes Ebook PDFDocument41 pagesUnderstanding Normal and Clinical Nutrition 11th Edition by Sharon Rady Rolfes Ebook PDFjames.auiles49297% (37)
- Microbiome-Directed Therapies - Past, Present, and Future Prebiotics and Probiotics in Digestive HealthDocument12 pagesMicrobiome-Directed Therapies - Past, Present, and Future Prebiotics and Probiotics in Digestive HealthcrackintheshatNo ratings yet
- Ulcerative ColitisDocument30 pagesUlcerative ColitisAndika SulistianNo ratings yet
- Crohns Disease PDFDocument40 pagesCrohns Disease PDFgcharala92No ratings yet
- Water Kefir Handbook - Water Kefir Recipes, Step-By-Step Instructions, Health Benefits and More (Water Kefir Recipes, Water Kefir For Beginners, Fermented Drinks, Fermented Foods Book 1) (PDFDrive)Document72 pagesWater Kefir Handbook - Water Kefir Recipes, Step-By-Step Instructions, Health Benefits and More (Water Kefir Recipes, Water Kefir For Beginners, Fermented Drinks, Fermented Foods Book 1) (PDFDrive)angelobuffaloNo ratings yet
- TG 9780199060870Document256 pagesTG 9780199060870HammadNo ratings yet
- IB HL Biology Notes: DigestionDocument9 pagesIB HL Biology Notes: DigestionayushfmNo ratings yet
- The Molecular Basis of Human CancerDocument868 pagesThe Molecular Basis of Human Cancerfamp_carrijo100% (8)
- Large Intestine Pattern Differentiation in Chinese MedicineDocument6 pagesLarge Intestine Pattern Differentiation in Chinese MedicineFinderlucian Lucian100% (1)
- Surgery Illustrated - Surgical Atlas: Mainz Pouch Continent Cutaneous DiversionDocument25 pagesSurgery Illustrated - Surgical Atlas: Mainz Pouch Continent Cutaneous DiversionAbdullah Bangwar100% (1)
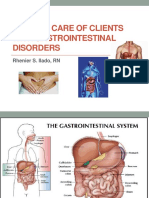
- Nursing of Clients With Gastrointestinal DisordersDocument228 pagesNursing of Clients With Gastrointestinal DisordersLane Mae Magpatoc Noerrot100% (1)
- Current Nutritional Therapies in Inflammatory Bowel Disease - Improving Clinical Remission Rates and Sustainability of Long-Term Dietary TherapiesDocument11 pagesCurrent Nutritional Therapies in Inflammatory Bowel Disease - Improving Clinical Remission Rates and Sustainability of Long-Term Dietary TherapiesAdriana SáNo ratings yet
- Diet-Microbiota Interactions in Inflammatory Bowel DiseaseDocument17 pagesDiet-Microbiota Interactions in Inflammatory Bowel DiseaseIsabella María GantivarNo ratings yet
- Nutrients 12 01423Document14 pagesNutrients 12 01423Nejc KovačNo ratings yet
- The Role of The FODMAP Diet in IBSDocument24 pagesThe Role of The FODMAP Diet in IBSMércia FiuzaNo ratings yet
- Nutrients 15 01351 v2Document12 pagesNutrients 15 01351 v2Diana DionisioNo ratings yet
- Dietary Fibre IBDDocument22 pagesDietary Fibre IBDErzsébet SoltészNo ratings yet
- Cells: Bile Acids: Key Players in Inflammatory Bowel Diseases?Document10 pagesCells: Bile Acids: Key Players in Inflammatory Bowel Diseases?Yaumil ChoiriNo ratings yet
- 2017 Nutrition Workshop ReportDocument6 pages2017 Nutrition Workshop ReportAmeliaNo ratings yet
- Pro y PrebioticosDocument21 pagesPro y PrebioticosIvan CLNo ratings yet
- From Discomfort To Delight Diets Impact On IBS DDocument10 pagesFrom Discomfort To Delight Diets Impact On IBS DAthenaeum Scientific PublishersNo ratings yet
- Short-Chain Fatty Acids As A Link Between Diet and Cardiometabolic Risk - A Narrative Review - PMCDocument25 pagesShort-Chain Fatty Acids As A Link Between Diet and Cardiometabolic Risk - A Narrative Review - PMCgabyzeragNo ratings yet
- Celiac DiseaseDocument19 pagesCeliac Disease35432No ratings yet
- Impact of Intensive Lifestyle Intervention On Gut Microbiota Composition in Type 2 Diabetes A Post-Hoc Analysis of A Randomized Clinical TrialDocument16 pagesImpact of Intensive Lifestyle Intervention On Gut Microbiota Composition in Type 2 Diabetes A Post-Hoc Analysis of A Randomized Clinical Trialvitoria.cristinaa62No ratings yet
- Diaz Gomis CN HealthydietaryDocument9 pagesDiaz Gomis CN HealthydietaryNoemy RodriguezNo ratings yet
- Campmans-Kuijpers 2021 IBDDocument34 pagesCampmans-Kuijpers 2021 IBDNovita ApramadhaNo ratings yet
- Research Proposal Final ProjectDocument16 pagesResearch Proposal Final Projectapi-534383603No ratings yet
- 3 - You Are What You Eat, Diet, HealthDocument22 pages3 - You Are What You Eat, Diet, HealthPaula CantalapiedraNo ratings yet
- BMJ 2023 075180fullDocument6 pagesBMJ 2023 075180fullPatyNo ratings yet
- Nutrients 13 00817 v2Document23 pagesNutrients 13 00817 v2Veronica Delgado QuinteroNo ratings yet
- Diabetes Obesity Metabolism - 2018 - Lee - Gut Microbiota and Obesity An Opportunity To Alter Obesity Through FaecalDocument12 pagesDiabetes Obesity Metabolism - 2018 - Lee - Gut Microbiota and Obesity An Opportunity To Alter Obesity Through FaecalNoemy RodriguezNo ratings yet
- Prebiotic Inulin-Type Fructans and Galacto-Oligosaccharides: de Finition, Specificity, Function, and Application in Gastrointestinal DisordersDocument5 pagesPrebiotic Inulin-Type Fructans and Galacto-Oligosaccharides: de Finition, Specificity, Function, and Application in Gastrointestinal DisordersGisela García PeñaNo ratings yet
- Food Fads: The Keto Diet and The Gut: Cause For Concern?Document2 pagesFood Fads: The Keto Diet and The Gut: Cause For Concern?Bogdan LeonardNo ratings yet
- Fnut 09 741630Document10 pagesFnut 09 741630gabrielavk95No ratings yet
- Gut Microbiota Modulation As A Possible Mediating Mechanism For Fasting-Induced Alleviation of Metabolic Complications: A Systematic ReviewDocument17 pagesGut Microbiota Modulation As A Possible Mediating Mechanism For Fasting-Induced Alleviation of Metabolic Complications: A Systematic ReviewalegmachadoNo ratings yet
- Xie PDocument19 pagesXie PSreeja CherukuruNo ratings yet
- NutrientsDocument13 pagesNutrientsDyane VatriciaNo ratings yet
- A Newly Developed Synbiotic Yogurt PreveDocument16 pagesA Newly Developed Synbiotic Yogurt PreveEvangelina RivarolaNo ratings yet
- Nutrients 14 02469 v2Document21 pagesNutrients 14 02469 v2Roxana CristeaNo ratings yet
- Metabolic Syndrome and Synbiotic Targeting of The Gut MicrobiomeDocument10 pagesMetabolic Syndrome and Synbiotic Targeting of The Gut Microbiomealpaca parcaNo ratings yet
- Polysaccharides From Natural Resources Exhibit Great Potential in The Treatment of Ulcerative Colitis - A ReviewDocument12 pagesPolysaccharides From Natural Resources Exhibit Great Potential in The Treatment of Ulcerative Colitis - A ReviewMartinaNo ratings yet
- Pre 2Document6 pagesPre 2ruNo ratings yet
- Art 2Document6 pagesArt 2Irene Ramirez ChavesNo ratings yet
- SCD ResearchDocument8 pagesSCD ResearchSharone BooisNo ratings yet
- Modulation of The Gastrointestinal Microbiome With Nondigestible Fermentable Carbohydrates To Improve Human HealthDocument35 pagesModulation of The Gastrointestinal Microbiome With Nondigestible Fermentable Carbohydrates To Improve Human HealthslothwonNo ratings yet
- Microorganisms 10 00452 v2Document15 pagesMicroorganisms 10 00452 v2V.EricSson BaPtisTaNo ratings yet
- Nutrients 12 01921 PDFDocument26 pagesNutrients 12 01921 PDFdianamsaucedoNo ratings yet
- Binge-Eating Disorder-CircadianDocument6 pagesBinge-Eating Disorder-CircadianMafe CordobaNo ratings yet
- PIIS2475299124000118Document18 pagesPIIS2475299124000118Reeds LaurelNo ratings yet
- Nutrients: Vegan Diet and The Gut Microbiota Composition in Healthy AdultsDocument14 pagesNutrients: Vegan Diet and The Gut Microbiota Composition in Healthy AdultsMiguel HuangNo ratings yet
- Articulo ProbioticosDocument13 pagesArticulo ProbioticososcarandresNo ratings yet
- 1 s2.0 S1521691813000619 Main Obezitatea Si MicrobiotaDocument11 pages1 s2.0 S1521691813000619 Main Obezitatea Si MicrobiotaLUTICHIEVICI NATALIANo ratings yet
- The Gut Microbiome As A Target For IBD Treatment - Are We There YetDocument12 pagesThe Gut Microbiome As A Target For IBD Treatment - Are We There YetMartinaNo ratings yet
- Gut Microbiota and Sex HormonesDocument21 pagesGut Microbiota and Sex HormonesGabriela DoniNo ratings yet
- Gizi Pada IBDDocument15 pagesGizi Pada IBDNovita ApramadhaNo ratings yet
- Nihms 833860Document20 pagesNihms 833860yabaNo ratings yet
- Enfermedad Inflamatoria IntestinalDocument10 pagesEnfermedad Inflamatoria IntestinalRebeca EspinoNo ratings yet
- OtimoDocument9 pagesOtimoRenata RothenNo ratings yet
- Final PaperDocument25 pagesFinal Paperapi-490571442No ratings yet
- Áteresztő Bél Sy És DMDocument12 pagesÁteresztő Bél Sy És DMDaniella FülöpNo ratings yet
- Nutrients: Can Gut Microbiota Composition Predict Response To Dietary Treatments?Document15 pagesNutrients: Can Gut Microbiota Composition Predict Response To Dietary Treatments?Meisya Nur'ainiNo ratings yet
- Nutrition, The Gut Microbiome and The Metabolic SyndromeDocument14 pagesNutrition, The Gut Microbiome and The Metabolic SyndromeM. Tasrif MansurNo ratings yet
- Bariatric Surgery in Obesity Efects On GutDocument27 pagesBariatric Surgery in Obesity Efects On GutMaria Bernardino CorrêaNo ratings yet
- Review: Stephanie Jew, Suhad S. Abumweis, and Peter J.H. JonesDocument10 pagesReview: Stephanie Jew, Suhad S. Abumweis, and Peter J.H. JonesJosé OrtízNo ratings yet
- Dietary Plants, Gut Microbiota, and Obesity. Effects and Mechanisms-DikonversiDocument18 pagesDietary Plants, Gut Microbiota, and Obesity. Effects and Mechanisms-DikonversiIndri Dewi ListianiNo ratings yet
- Reviews: You Are What You Eat: Diet, Health and The Gut MicrobiotaDocument22 pagesReviews: You Are What You Eat: Diet, Health and The Gut MicrobiotaHarry YucraNo ratings yet
- Nutrients 13 00526Document16 pagesNutrients 13 00526yabaNo ratings yet
- Microbial Therapeutics in Neurocognitive and Psychiatric DisordersDocument21 pagesMicrobial Therapeutics in Neurocognitive and Psychiatric Disorderssamira6alvarado6zebaNo ratings yet
- Dietary Fibre Functionality in Food and Nutraceuticals: From Plant to GutFrom EverandDietary Fibre Functionality in Food and Nutraceuticals: From Plant to GutFarah HosseinianNo ratings yet
- Exploring Novel Colon-Targeting Antihistaminic Prodrug For ColitisDocument11 pagesExploring Novel Colon-Targeting Antihistaminic Prodrug For ColitisNejc KovačNo ratings yet
- Frontiers in Bioscience S4, 226-239, January 1, 2012Document14 pagesFrontiers in Bioscience S4, 226-239, January 1, 2012Nejc KovačNo ratings yet
- Major Advances in The Development of Histamine H Receptor LigandsDocument9 pagesMajor Advances in The Development of Histamine H Receptor LigandsNejc KovačNo ratings yet
- HHS Public Access: The Multifaceted Mast Cell in Inflammatory Bowel DiseaseDocument29 pagesHHS Public Access: The Multifaceted Mast Cell in Inflammatory Bowel DiseaseNejc KovačNo ratings yet
- FFFFFDocument22 pagesFFFFFNejc KovačNo ratings yet
- F 5Document16 pagesF 5Nejc KovačNo ratings yet
- Effect of C-Kit Ligand, Factor, Release by Human InflammatoryDocument11 pagesEffect of C-Kit Ligand, Factor, Release by Human InflammatoryNejc KovačNo ratings yet
- The Effect of Acute Psychologic Stress On Systemic and Rectal Mucosal Measures of in Ammation in Ulcerative ColitisDocument10 pagesThe Effect of Acute Psychologic Stress On Systemic and Rectal Mucosal Measures of in Ammation in Ulcerative ColitisNejc KovačNo ratings yet
- F 1Document11 pagesF 1Nejc KovačNo ratings yet
- Regulation of The Immune Response and Inflammation by Histamine and Histamine ReceptorsDocument10 pagesRegulation of The Immune Response and Inflammation by Histamine and Histamine ReceptorsNejc KovačNo ratings yet
- Secretagogue Response of Goblet Cells and Columnar Cells in Human Colonic CryptsDocument22 pagesSecretagogue Response of Goblet Cells and Columnar Cells in Human Colonic CryptsNejc KovačNo ratings yet
- NIH Public Access: Author ManuscriptDocument9 pagesNIH Public Access: Author ManuscriptNejc KovačNo ratings yet
- Release of Mast Cell Tryptase From Human Colorectal Mucosa in Inflammatory Bowel DiseaseDocument7 pagesRelease of Mast Cell Tryptase From Human Colorectal Mucosa in Inflammatory Bowel DiseaseNejc KovačNo ratings yet
- Evidence For Mast Cell Activation in Collagenous Colitis: Inflammation ResearchDocument2 pagesEvidence For Mast Cell Activation in Collagenous Colitis: Inflammation ResearchNejc KovačNo ratings yet
- Effects of Amine Oxidases in Allergic and Histamine-Mediated ConditionsDocument35 pagesEffects of Amine Oxidases in Allergic and Histamine-Mediated ConditionsNejc KovačNo ratings yet
- DDDDDocument11 pagesDDDDNejc KovačNo ratings yet
- Version of RecordDocument16 pagesVersion of RecordNejc KovačNo ratings yet
- Histamine H2 Receptor-Mediated Suppression of Intestinal Inflammation by Probiotic Lactobacillus ReuteriDocument12 pagesHistamine H2 Receptor-Mediated Suppression of Intestinal Inflammation by Probiotic Lactobacillus ReuteriNejc KovačNo ratings yet
- HHS Public AccessDocument23 pagesHHS Public AccessNejc KovačNo ratings yet
- D 6Document8 pagesD 6Nejc KovačNo ratings yet
- Enhancement of Human Intestinal Mast Cell Mediator Release in Active Ulcerative ColitisDocument6 pagesEnhancement of Human Intestinal Mast Cell Mediator Release in Active Ulcerative ColitisNejc KovačNo ratings yet
- Histamine, Histamine Intoxication and Intolerance: Allergologia Et ImmunopathologiaDocument9 pagesHistamine, Histamine Intoxication and Intolerance: Allergologia Et ImmunopathologiaNejc KovačNo ratings yet
- Distinct Roles of Histamine H1 and H2 Receptor Signaling Pathways in Inflammation-Associated Colonic TumorigenesisDocument35 pagesDistinct Roles of Histamine H1 and H2 Receptor Signaling Pathways in Inflammation-Associated Colonic TumorigenesisNejc KovačNo ratings yet
- BBBBDocument18 pagesBBBBNejc KovačNo ratings yet
- Histamine H Receptors in The Gastrointestinal Tract: ReviewDocument14 pagesHistamine H Receptors in The Gastrointestinal Tract: ReviewNejc KovačNo ratings yet
- Histamine: An Undercover Agent in Multiple Rare Diseases?: J. Cell. Mol. Med. Vol 16, No 9, 2012 Pp. 1947-1960Document14 pagesHistamine: An Undercover Agent in Multiple Rare Diseases?: J. Cell. Mol. Med. Vol 16, No 9, 2012 Pp. 1947-1960Nejc KovačNo ratings yet
- Aaaaa 5Document15 pagesAaaaa 5Nejc KovačNo ratings yet
- Gut Microbe Emediated Suppression of in Ammation-Associated Colon Carcinogenesis by Luminal Histamine ProductionDocument14 pagesGut Microbe Emediated Suppression of in Ammation-Associated Colon Carcinogenesis by Luminal Histamine ProductionNejc KovačNo ratings yet
- The Taxonomic Distribution of Histamine-Secreting Bacteria in The Human Gut MicrobiomeDocument11 pagesThe Taxonomic Distribution of Histamine-Secreting Bacteria in The Human Gut MicrobiomeNejc KovačNo ratings yet
- Key Role of Mast Cells and Their Major Secretory Products in Inflammatory Bowel DiseaseDocument10 pagesKey Role of Mast Cells and Their Major Secretory Products in Inflammatory Bowel DiseaseNejc KovačNo ratings yet
- Theresa Porrett, Anthony McGrath - Stoma Care (Essential Clinical Skills For Nurses) (2005)Document196 pagesTheresa Porrett, Anthony McGrath - Stoma Care (Essential Clinical Skills For Nurses) (2005)Drogo MejezelNo ratings yet
- Acupressure Points For Constipation and Diarrhrea ReliefDocument5 pagesAcupressure Points For Constipation and Diarrhrea ReliefConstantin-Mihail PopescuNo ratings yet
- What Causes Diarrhea After EatingDocument6 pagesWhat Causes Diarrhea After Eatingyushendra putraNo ratings yet
- Non Traumatic Acute Abdomen in Pediatric Patients: US Imaging With Clinical and Surgical Correlation-A Case-Base...Document39 pagesNon Traumatic Acute Abdomen in Pediatric Patients: US Imaging With Clinical and Surgical Correlation-A Case-Base...Vina RatnasariNo ratings yet
- Ns4 m2 U2 PresentationDocument16 pagesNs4 m2 U2 PresentationYulia ChernykhNo ratings yet
- This Study Resource Was: Digestive SystemDocument4 pagesThis Study Resource Was: Digestive SystemMark Jerome CifraNo ratings yet
- Intricate Process of DigestionDocument1 pageIntricate Process of DigestionARC THE BUMPNo ratings yet
- LSM Grade 4 Science 1st Trim Exam SY 2010 - 2011Document10 pagesLSM Grade 4 Science 1st Trim Exam SY 2010 - 2011Mauie Flores100% (1)
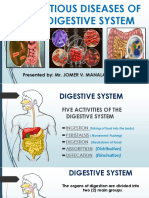
- Communicable Diseases of The GITDocument31 pagesCommunicable Diseases of The GITNicole Anne TungolNo ratings yet
- Gen Ana and PhysioDocument360 pagesGen Ana and Physioanadelavega96No ratings yet
- Human Biology 13Th Edition Mader Test Bank Full Chapter PDFDocument56 pagesHuman Biology 13Th Edition Mader Test Bank Full Chapter PDFninhdermotc1u100% (9)
- Structural Organisation in AnimalsDocument9 pagesStructural Organisation in AnimalsPrincipal EscetNo ratings yet
- Crohns Case StudyDocument16 pagesCrohns Case Studyapi-375211204No ratings yet
- CARBOHYDRATE ABSORPTION (Ref)Document22 pagesCARBOHYDRATE ABSORPTION (Ref)NEERAJA S RAJNo ratings yet
- The Ordog Effect 3Document8 pagesThe Ordog Effect 3Jw SqNo ratings yet
- Development of The Alimentary System - IDocument27 pagesDevelopment of The Alimentary System - Iمعاذ القحطانيNo ratings yet
- Parte 2 Complete Reflexology For Live (Inglés) Autor Barbara y Kevin Kunz-262-289Document28 pagesParte 2 Complete Reflexology For Live (Inglés) Autor Barbara y Kevin Kunz-262-289Hombres Alto ValorNo ratings yet
- Sci F2 T2 (E)Document7 pagesSci F2 T2 (E)Nad RamliNo ratings yet