You might also like
- Normal Impulse Conduction: Sinoatrial Node AV Node Bundle of His Bundle Branches Purkinje FibersDocument73 pagesNormal Impulse Conduction: Sinoatrial Node AV Node Bundle of His Bundle Branches Purkinje FibersIndra ToshiwayNo ratings yet
- Kisi Kisi EndoDocument2 pagesKisi Kisi EndoIndra ToshiwayNo ratings yet
- Antihypertensive Drug Classes & MechanismsDocument23 pagesAntihypertensive Drug Classes & MechanismsIndra ToshiwayNo ratings yet
- Antihypertensive AgentDocument23 pagesAntihypertensive AgentIndra ToshiwayNo ratings yet
- Kolesterol Blok CvsDocument82 pagesKolesterol Blok CvsFadilah NSNo ratings yet
- Normal Impulse Conduction and ECG InterpretationDocument73 pagesNormal Impulse Conduction and ECG InterpretationIndra ToshiwayNo ratings yet
- Xeno BioticDocument54 pagesXeno BioticIndra ToshiwayNo ratings yet
- Mumps and Measles Virus 2010Document31 pagesMumps and Measles Virus 2010Indra ToshiwayNo ratings yet
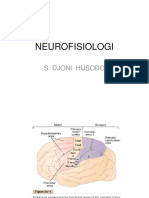
- Neurofisiologi: S Djoni HusodoDocument7 pagesNeurofisiologi: S Djoni HusodoIndra ToshiwayNo ratings yet
- Reovirus Rotavirus NewDocument19 pagesReovirus Rotavirus NewMuhammad Deyanta HafidzNo ratings yet
- Immunology To InfectionDocument47 pagesImmunology To InfectionIndra ToshiwayNo ratings yet
- Kuliah Cacing WestyDocument63 pagesKuliah Cacing WestyIndra ToshiwayNo ratings yet
- Picornavirus XDocument26 pagesPicornavirus XIndra ToshiwayNo ratings yet
- Lymphatic Circulation Revised 2011Document34 pagesLymphatic Circulation Revised 2011Indra ToshiwayNo ratings yet
- Immunology To InfectionDocument47 pagesImmunology To InfectionIndra ToshiwayNo ratings yet
- Majority of amino acids used for de novo protein synthesis derives from protein degradation (80Document47 pagesMajority of amino acids used for de novo protein synthesis derives from protein degradation (80Indra ToshiwayNo ratings yet
- AutacoidDocument36 pagesAutacoidIndra ToshiwayNo ratings yet
- AnticoagulantsDocument20 pagesAnticoagulantsIndra ToshiwayNo ratings yet
- AnemiaDocument33 pagesAnemiaIndra ToshiwayNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Structure and Classes of Immunoglobulins (IgDocument5 pagesStructure and Classes of Immunoglobulins (IgsajjadNo ratings yet
- National Immunization Schedule (NIS) : For Infants, Children and Pregnant WomenDocument13 pagesNational Immunization Schedule (NIS) : For Infants, Children and Pregnant WomenPrabir Kumar Chatterjee100% (1)
- Immunology Mcqs-III (Gate Helpline)Document4 pagesImmunology Mcqs-III (Gate Helpline)Santhosh Kalash75% (4)
- Allergic Rhinitis in Children: Diagnosis and TreatmentDocument39 pagesAllergic Rhinitis in Children: Diagnosis and Treatmentrinajacky100% (2)
- Bioterrorism: BY: - Bijay Kumar Mahato BPH 2 Semester Padmashree School of Public HealthDocument26 pagesBioterrorism: BY: - Bijay Kumar Mahato BPH 2 Semester Padmashree School of Public HealthBijay Kumar MahatoNo ratings yet
- Detect Dengue NS1 Antigen, IgG & IgM (RapidDocument1 pageDetect Dengue NS1 Antigen, IgG & IgM (RapidSure NavyasriNo ratings yet
- Communicable Disease Nursing: Hosts, Microbes and ImmunityDocument421 pagesCommunicable Disease Nursing: Hosts, Microbes and ImmunityJoric Magusara91% (11)
- RBC Haematology MCQsDocument29 pagesRBC Haematology MCQsHabib UllahNo ratings yet
- Articulo 4.Document11 pagesArticulo 4.Yajaira MenesesNo ratings yet
- Autoimmune Hepatitis HandoutDocument2 pagesAutoimmune Hepatitis HandoutClarisse SantosNo ratings yet
- Students ProfileDocument1 pageStudents ProfileHuegene Andreu GabiNo ratings yet
- MODULE 4 (Basics of STI, HIV & AIDS)Document45 pagesMODULE 4 (Basics of STI, HIV & AIDS)ninafatima allamNo ratings yet
- Session 5 Part 1Document10 pagesSession 5 Part 1Antonette Alexis AnchetaNo ratings yet
- Case 1: Group 4Document26 pagesCase 1: Group 4John Joshua Lacson MedicineNo ratings yet
- Chapter 22: Immunizations Garzon Maaks: Burns' Pediatric Primary Care, 7th EditionDocument2 pagesChapter 22: Immunizations Garzon Maaks: Burns' Pediatric Primary Care, 7th EditionHelen UgochukwuNo ratings yet
- Hospital Antibiotic Policy: ST Helens and Knowsley Hospitals TrustDocument72 pagesHospital Antibiotic Policy: ST Helens and Knowsley Hospitals TrustDimas RfNo ratings yet
- Antibody IdentificationDocument27 pagesAntibody Identificationhamaada100% (1)
- 8th April 2012 IssueDocument4 pages8th April 2012 IssuebcmbtNo ratings yet
- The Man With The Golden Blood - MosaicDocument11 pagesThe Man With The Golden Blood - MosaicagxzNo ratings yet
- 2nd Half Microbiology For Allied Health StudentsDocument404 pages2nd Half Microbiology For Allied Health StudentsFjornNo ratings yet
- Oral Tolerance:: The Response of The Intestinal Mucosa To Dietary AntigensDocument218 pagesOral Tolerance:: The Response of The Intestinal Mucosa To Dietary AntigensammarkochiNo ratings yet
- Inflammation: InfectionDocument11 pagesInflammation: InfectionRenBautistaNo ratings yet
- BDS FINAL YEAR PERIODONTICS CHAPTERSDocument7 pagesBDS FINAL YEAR PERIODONTICS CHAPTERSakash_81087No ratings yet
- How Pfizer Delivered A Covid Vaccine in Record Time - WSJDocument17 pagesHow Pfizer Delivered A Covid Vaccine in Record Time - WSJvibesjayNo ratings yet
- Spinal MeningitisDocument7 pagesSpinal Meningitisapi-282958026No ratings yet
- Scabies Signs, Symptoms & TreatmentDocument4 pagesScabies Signs, Symptoms & TreatmentFerreze AnnNo ratings yet
- Patogenia SAFDocument39 pagesPatogenia SAFTra21798siempreNo ratings yet
- Martha Mamora (Adisseo) - Munas SCI 2022Document14 pagesMartha Mamora (Adisseo) - Munas SCI 2022ifaNo ratings yet
- Infection Control, Safety First Aid and Personal WellnessDocument9 pagesInfection Control, Safety First Aid and Personal WellnessJacinta MalamionNo ratings yet
- Rheumatic Heart DiseaseDocument25 pagesRheumatic Heart DiseaseMag5No ratings yet