You might also like
- Chapter 19Document42 pagesChapter 19Dr. Manish KumarNo ratings yet
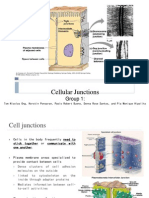
- Cell JunctionDocument63 pagesCell JunctionsphuyalNo ratings yet
- Cell JunctionsDocument56 pagesCell JunctionsDrGagandeep Singh100% (3)
- Aberrant - Brainwaves (Unpublished)Document55 pagesAberrant - Brainwaves (Unpublished)Stuart Goheen100% (2)
- Cell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemDocument70 pagesCell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemGovinda BiswasNo ratings yet
- BM 8-9 OncogenesisDocument139 pagesBM 8-9 OncogenesisabdullahshiddiqadamNo ratings yet
- of Cell Adhesion and Cell JunctionsDocument50 pagesof Cell Adhesion and Cell Junctionsayad ghawiNo ratings yet
- Gene Regulation: Mrs. Ofelia Solano SaludarDocument39 pagesGene Regulation: Mrs. Ofelia Solano SaludarmskikiNo ratings yet
- Cell JunctionsDocument40 pagesCell JunctionsAhmad MursyidNo ratings yet
- Cell Cycle and Mitosis 2017Document26 pagesCell Cycle and Mitosis 2017Jodi Davis0% (1)
- Crispr Case StudyDocument29 pagesCrispr Case Studyapi-330846676No ratings yet
- Cell JunctionsDocument34 pagesCell JunctionsDrAmit Gaba Mds100% (1)
- Mammalian Histology AssignmentDocument9 pagesMammalian Histology AssignmentSana Sultana100% (1)
- BOOK Gene TherapyDocument210 pagesBOOK Gene TherapyCarla GomesNo ratings yet
- Cell JunctionsDocument30 pagesCell JunctionsNavodit GoelNo ratings yet
- Keratinisation and CornificationDocument38 pagesKeratinisation and Cornificationlizamjen100% (3)
- CellDocument47 pagesCellAnami NizamNo ratings yet
- En Con Celladhesion 20120407Document17 pagesEn Con Celladhesion 20120407CLPHtheoryNo ratings yet
- Cell Adhesion and Cell Adhesion MoleculesDocument7 pagesCell Adhesion and Cell Adhesion MoleculesLemon GundersonNo ratings yet
- CelladhesionDocument51 pagesCelladhesionNina Difla MuflikhahNo ratings yet
- Epithelium Surface SpecializationsDocument41 pagesEpithelium Surface SpecializationsMitzel SapaloNo ratings yet
- Cell-to-Cell and Cell-to-Matrix Adhesions: Fibronectin?Document46 pagesCell-to-Cell and Cell-to-Matrix Adhesions: Fibronectin?MinarsihNo ratings yet
- Class 19 CellBiology MB 2023 2024Document20 pagesClass 19 CellBiology MB 2023 2024André MatosNo ratings yet
- Integrasi Sel Dalam Jaringan (Kuliah) - 2 - 2Document125 pagesIntegrasi Sel Dalam Jaringan (Kuliah) - 2 - 2komang nickoNo ratings yet
- Celladhesion Epithel KissADocument51 pagesCelladhesion Epithel KissAamdusuf33No ratings yet
- CellCell Webdez13Document37 pagesCellCell Webdez13viralnanobio_4150420No ratings yet
- Cell Surface receptorsMCBA P2 TDocument16 pagesCell Surface receptorsMCBA P2 TMatende husseinNo ratings yet
- Partoza, J WS Cel - MolDocument6 pagesPartoza, J WS Cel - MolJenny PartozaNo ratings yet
- Biochemistry Extracellular MatrixDocument5 pagesBiochemistry Extracellular MatrixbrianhelbigNo ratings yet
- Cadherin: Cadherins (Named For "Calcium-DependentDocument10 pagesCadherin: Cadherins (Named For "Calcium-Dependentfelipe smithNo ratings yet
- Bookmark This Page: Cadherins Mucin-Like Cams IntegrinsDocument38 pagesBookmark This Page: Cadherins Mucin-Like Cams IntegrinsAndreea SpiridonNo ratings yet
- Physiological Processes Where Cell Adhesion Is ImportantDocument42 pagesPhysiological Processes Where Cell Adhesion Is ImportantJT92No ratings yet
- Cell Adhesion MoleculesDocument14 pagesCell Adhesion MoleculesSecret Agent100% (1)
- Biological Considerations 3.1 The Culture EnvironmentDocument4 pagesBiological Considerations 3.1 The Culture EnvironmentPaavni SharmaNo ratings yet
- Extracellular Matrix: Mrs. Ofelia Solano SaludarDocument29 pagesExtracellular Matrix: Mrs. Ofelia Solano SaludarmskikiNo ratings yet
- 06lect ECM Summary2017Document5 pages06lect ECM Summary2017Dan ChoiNo ratings yet
- Cell AdhesinDocument2 pagesCell Adhesinapi-3812274No ratings yet
- Chapter 4-Jnu Biomembranes-1 - 複本Document48 pagesChapter 4-Jnu Biomembranes-1 - 複本Wai Kwong ChiuNo ratings yet
- Cell AdhesionDocument13 pagesCell Adhesionaredhel ar-feinielNo ratings yet
- Cell CompartmentalizationDocument36 pagesCell Compartmentalizationanuj2907No ratings yet
- 3 Cytology Membran JunctionDocument48 pages3 Cytology Membran JunctionSamNo ratings yet
- Organelos CelularesDocument47 pagesOrganelos CelularesYOSUAR ANTONIO SALGADO HERNANDEZNo ratings yet
- Extracell MatrixDocument14 pagesExtracell MatrixValine Cysteine MethionineNo ratings yet
- ANAT309 2.cell 1Document35 pagesANAT309 2.cell 1Pushparaj ShettyNo ratings yet
- Cell Junctions, Gap Adhesions, and ExtracellularDocument58 pagesCell Junctions, Gap Adhesions, and ExtracellularFabian Christine MaeNo ratings yet
- Cell Bio 3 Cell Junctions 2021 PDFDocument38 pagesCell Bio 3 Cell Junctions 2021 PDFaboodmr8nblahNo ratings yet
- N-Linked Protein Glycosylation Occurs in All Three Life-Form Domains: Eukaryotes, Bacteria, andDocument9 pagesN-Linked Protein Glycosylation Occurs in All Three Life-Form Domains: Eukaryotes, Bacteria, andHeisenbergNo ratings yet
- Static and Dynamic Interactions Between Endothelium and Circulating Cells in CancerDocument25 pagesStatic and Dynamic Interactions Between Endothelium and Circulating Cells in CancerderNo ratings yet
- AdhesionDocument4 pagesAdhesionnuman88No ratings yet
- Unit 11 Cell AdesionDocument15 pagesUnit 11 Cell AdesionSarah PavuNo ratings yet
- JunctionsDocument96 pagesJunctionsAnyone Can CookNo ratings yet
- Lectures 11-13 NotesDocument10 pagesLectures 11-13 Notesapi-3721438No ratings yet
- Cell JunctionsDocument29 pagesCell JunctionsMira Cindy IINo ratings yet
- While Red Blood Cells Are Carried Away at High Velocity by A Strong Blood FlowDocument2 pagesWhile Red Blood Cells Are Carried Away at High Velocity by A Strong Blood Flowberfin şakNo ratings yet
- Scie-14 Interaction Between Cells and Their EnvironmentDocument13 pagesScie-14 Interaction Between Cells and Their EnvironmentAliah MosqueraNo ratings yet
- Cell JunctionsDocument24 pagesCell JunctionsNajlaNo ratings yet
- Cell Junctions: Mrs. Ofelia Solano SaludarDocument19 pagesCell Junctions: Mrs. Ofelia Solano SaludarmskikiNo ratings yet
- 8 Non-Muscle Actin, IFs, Adhesion-2Document35 pages8 Non-Muscle Actin, IFs, Adhesion-2zj5bnxbymzNo ratings yet
- Brooker2014 COHESINAS PDFDocument38 pagesBrooker2014 COHESINAS PDFMary MontoyaNo ratings yet
- Selectins: Dr. Muhammad Ali Faridi Dr. Arathy JosephDocument17 pagesSelectins: Dr. Muhammad Ali Faridi Dr. Arathy JosephAli FaridiNo ratings yet
- Cellular JunctionsDocument25 pagesCellular JunctionsDiana Grace Pile0% (1)
- Adhesion Signalling Complexes: PrimerDocument5 pagesAdhesion Signalling Complexes: PrimerdupuytrenNo ratings yet
- 3 B Cytology-Membrane StructuresDocument19 pages3 B Cytology-Membrane Structuresm.mansoor3377No ratings yet
- CT Proper ComponentsDocument55 pagesCT Proper ComponentsMitzel SapaloNo ratings yet
- Matriks EkstraselulerDocument75 pagesMatriks EkstraselulerRenaldy PamungkasNo ratings yet
- Mrs. Ofelia Solano Saludar: Department of Natural Sciences University of St. La Salle Bacolod CityDocument57 pagesMrs. Ofelia Solano Saludar: Department of Natural Sciences University of St. La Salle Bacolod CitymskikiNo ratings yet
- Translation: Mrs. Ofelia Solano SaludarDocument28 pagesTranslation: Mrs. Ofelia Solano SaludarmskikiNo ratings yet
- Mrs. Ofelia Solano Saludar: Department of Natural Sciences University of St. La Salle Bacolod CityDocument55 pagesMrs. Ofelia Solano Saludar: Department of Natural Sciences University of St. La Salle Bacolod CitymskikiNo ratings yet
- Protein Structure, Targeting and SortingDocument28 pagesProtein Structure, Targeting and SortingmskikiNo ratings yet
- Mutation and Dna Repair MechanismsDocument46 pagesMutation and Dna Repair MechanismsmskikiNo ratings yet
- Extracellular Matrix: Mrs. Ofelia Solano SaludarDocument29 pagesExtracellular Matrix: Mrs. Ofelia Solano SaludarmskikiNo ratings yet
- Anatomy of A GeneDocument33 pagesAnatomy of A GenemskikiNo ratings yet
- Mrs. Ofelia Solano Saludar: Department of Natural Sciences University of St. La Salle Bacolod CityDocument33 pagesMrs. Ofelia Solano Saludar: Department of Natural Sciences University of St. La Salle Bacolod CitymskikiNo ratings yet
- Ligands and Signal Trans Duct IonDocument36 pagesLigands and Signal Trans Duct IonmskikiNo ratings yet
- Cell Junctions: Mrs. Ofelia Solano SaludarDocument19 pagesCell Junctions: Mrs. Ofelia Solano SaludarmskikiNo ratings yet
- Cell Biology: An Introduction: Mrs. Ofelia Solano SaludarDocument29 pagesCell Biology: An Introduction: Mrs. Ofelia Solano SaludarmskikiNo ratings yet
- Visualizing Nucleic AcidsDocument43 pagesVisualizing Nucleic AcidsmskikiNo ratings yet
- Visualizing Molecules in Living CellsDocument42 pagesVisualizing Molecules in Living CellsmskikiNo ratings yet
- Methods of Investigati NG Cells: Mrs. Ofelia Solano SaludarDocument35 pagesMethods of Investigati NG Cells: Mrs. Ofelia Solano SaludarmskikiNo ratings yet
- CARBOHYDRATES With Case StudyDocument1 pageCARBOHYDRATES With Case StudyAweGooseTreeNo ratings yet
- Pharmaceuticals & Medical Supplies: Amarila Malik Professor, Faculty of Pharmacy - Universitas IndonesiaDocument22 pagesPharmaceuticals & Medical Supplies: Amarila Malik Professor, Faculty of Pharmacy - Universitas IndonesiaAngga AnugrawanNo ratings yet
- Keanekaragaman Bakteri Penghasil EksopolisakaridaDocument8 pagesKeanekaragaman Bakteri Penghasil EksopolisakaridaMartinus GintingNo ratings yet
- Biochemistry Concepts and Connections 1St Edition Appling Test Bank Full Chapter PDFDocument27 pagesBiochemistry Concepts and Connections 1St Edition Appling Test Bank Full Chapter PDFElizabethRuizrxka100% (13)
- STPM Syllabus ProteinDocument6 pagesSTPM Syllabus ProteinOON XIN YI MoeNo ratings yet
- Original Pdfbiology Exploring The Diversity of Life 4th 1 3 by Peter Russell PDFDocument41 pagesOriginal Pdfbiology Exploring The Diversity of Life 4th 1 3 by Peter Russell PDFbonnie.eaton513100% (36)
- Stem Cells WordDocument6 pagesStem Cells Wordaaroncalebjeba70No ratings yet
- A Guided Tour of The Living CellDocument2 pagesA Guided Tour of The Living CellImam KharomainNo ratings yet
- Detection of Homozygous and Hemizygous Complete or Partial Exon Deletions by Whole-Exome SequencingDocument10 pagesDetection of Homozygous and Hemizygous Complete or Partial Exon Deletions by Whole-Exome SequencingMoundir AbderrahmaneNo ratings yet
- Research Paper On Plant Biotechnology PDFDocument6 pagesResearch Paper On Plant Biotechnology PDFfveec9sx100% (1)
- Immunization PPT - EditedDocument22 pagesImmunization PPT - EditedAlfred Mutua MutungaNo ratings yet
- Data Sharing Policy and Guidelines Jan 2021Document10 pagesData Sharing Policy and Guidelines Jan 2021Keren HilaryNo ratings yet
- Origin of Replication: Gram Negative Bacterial PlasmidsDocument12 pagesOrigin of Replication: Gram Negative Bacterial PlasmidsMagesh RamasamyNo ratings yet
- Ascorbate and Glutathione: Keeping Active Oxygen Under ControlDocument31 pagesAscorbate and Glutathione: Keeping Active Oxygen Under ControlPhilip Blair OngNo ratings yet
- Proteins, Lecture 5Document34 pagesProteins, Lecture 5xdawy2004xdNo ratings yet
- Elsevier 1 N08Document270 pagesElsevier 1 N08criticald0% (1)
- Chapter 12 DNA and RNA ReviewDocument6 pagesChapter 12 DNA and RNA ReviewAndrew WatsonNo ratings yet
- DNA Microarray Technology 1Document5 pagesDNA Microarray Technology 1NoorNo ratings yet
- 11 Introduction To Molecular Medicine PDFDocument11 pages11 Introduction To Molecular Medicine PDFErlinda NavalloNo ratings yet
- Recording Forms g1 g4 g7 ImmunizationDocument4 pagesRecording Forms g1 g4 g7 ImmunizationGirlie Harical GangawanNo ratings yet
- Chapter 05Document54 pagesChapter 05HasanNo ratings yet
- Full Download Test Bank For Biochemistry A Short Course 2nd Edition John L Tymoczko Isbn 10 1429283602 Isbn 13 9781429283601 PDF Full ChapterDocument36 pagesFull Download Test Bank For Biochemistry A Short Course 2nd Edition John L Tymoczko Isbn 10 1429283602 Isbn 13 9781429283601 PDF Full Chapteranotheraback.i39h100% (18)
- PESTLE Analysis of Pharma SectorDocument5 pagesPESTLE Analysis of Pharma Sectorshambhavi jhaNo ratings yet
- Plant Form and FunctionDocument43 pagesPlant Form and FunctionMae MejaresNo ratings yet
- Molecular Biology - WikipediaDocument9 pagesMolecular Biology - WikipediaLizbethNo ratings yet