You might also like
- Camp's Zoology by the Numbers: A comprehensive study guide in outline form for advanced biology courses, including AP, IB, DE, and college courses.From EverandCamp's Zoology by the Numbers: A comprehensive study guide in outline form for advanced biology courses, including AP, IB, DE, and college courses.No ratings yet
- Drosophila DevelopmentDocument63 pagesDrosophila DevelopmentabdurNo ratings yet
- Early Embryology, Fate Determination, and Patterning in DrosophilaDocument63 pagesEarly Embryology, Fate Determination, and Patterning in DrosophilaClarisse CalingasanNo ratings yet
- Genetic Analysis of Fruit Fly Embryogenesis Reveals Conserved Developmental MechanismsDocument60 pagesGenetic Analysis of Fruit Fly Embryogenesis Reveals Conserved Developmental MechanismsEnzo TapingNo ratings yet
- D MelanogasterDocument40 pagesD Melanogastersunifeb128075No ratings yet
- Pattern Formation in DrosophilaDocument17 pagesPattern Formation in DrosophilaAdwika DeoNo ratings yet
- Genetics of Axis Specification in DrosophilaDocument3 pagesGenetics of Axis Specification in DrosophilaJohnkevin NocheNo ratings yet
- Drosophila: 4 BIO-8 (Group 5)Document98 pagesDrosophila: 4 BIO-8 (Group 5)AudreyNo ratings yet
- BIO125ZusammenfassungLF 3Document43 pagesBIO125ZusammenfassungLF 399rnbccdr8No ratings yet
- Drosophila Embryogenesis: The Formation of Fruit Fly EmbryosDocument6 pagesDrosophila Embryogenesis: The Formation of Fruit Fly EmbryosVikram Pkb SharmaNo ratings yet
- Pattern Formation in DrosophilaDocument18 pagesPattern Formation in DrosophilacorashersNo ratings yet
- 1519024439M06AxisspecificationindrosophilaQuad1Document18 pages1519024439M06AxisspecificationindrosophilaQuad1Mahendra KatreNo ratings yet
- Cell Biology Ch21 Part 1 05222023 강의자료Document32 pagesCell Biology Ch21 Part 1 05222023 강의자료조형윤No ratings yet
- Genetics of Axis Specification in Drosophila Part 1 - Basics of Drosophila EmbryogenesisDocument28 pagesGenetics of Axis Specification in Drosophila Part 1 - Basics of Drosophila Embryogenesisrheen77No ratings yet
- CHAPTER 11 - Drosophila: Cleavage and BlastulaDocument5 pagesCHAPTER 11 - Drosophila: Cleavage and BlastulaAngela BalmesNo ratings yet
- Fate Maps: Course: B.Sc. (H) Zoology VI Semester Paper: Developmental Biology Faculty: Dr. Priya GoelDocument17 pagesFate Maps: Course: B.Sc. (H) Zoology VI Semester Paper: Developmental Biology Faculty: Dr. Priya GoelSusmita PalNo ratings yet
- Developmental Biology I: Classical Embryology and the Emergence of Organised StructuresDocument24 pagesDevelopmental Biology I: Classical Embryology and the Emergence of Organised StructuresSonali TayalNo ratings yet
- Zoo301lex2 4 Intros Alviar2017Document2 pagesZoo301lex2 4 Intros Alviar2017Mary Rose SantiagoNo ratings yet
- Animal Development: Cell Biology October 23, 2013Document16 pagesAnimal Development: Cell Biology October 23, 2013RustEdNo ratings yet
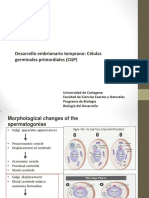
- Desarrollo Embrionario Temprano: Células Germinales Primordiales (CGP)Document30 pagesDesarrollo Embrionario Temprano: Células Germinales Primordiales (CGP)Yar CBNo ratings yet
- Cell Transport and Membrane StructureDocument6 pagesCell Transport and Membrane StructureGonzaga, Kyle P.No ratings yet
- Lec. 13 Drosophila 2 PDFDocument53 pagesLec. 13 Drosophila 2 PDFBasma RagabNo ratings yet
- EMBRYOLOGY Part1Document90 pagesEMBRYOLOGY Part1Daly DaliaNo ratings yet
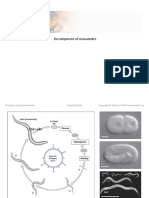
- Nematodes DevelopmentDocument31 pagesNematodes DevelopmentAbhisekh ChhatarNo ratings yet
- Molecular Biology of Reproductive ProcessesDocument53 pagesMolecular Biology of Reproductive ProcessesBhaskar GangulyNo ratings yet
- Week 2 Lecture (UCLA Ex)Document51 pagesWeek 2 Lecture (UCLA Ex)Soji AdimulaNo ratings yet
- MBB 142 LE1 Notes - Word VersionDocument72 pagesMBB 142 LE1 Notes - Word VersionWeaverly Colleen LeeNo ratings yet
- Development of Drosophila: Hand-Out CBT Chapter 2 Wolpert, 5 Edition March 2018Document15 pagesDevelopment of Drosophila: Hand-Out CBT Chapter 2 Wolpert, 5 Edition March 2018eze033No ratings yet
- Trans - Patten's Foundations of Embryology Chapters 1-7Document21 pagesTrans - Patten's Foundations of Embryology Chapters 1-7Lorenzo Daniel Antonio0% (1)
- Cell Specification MechanismsDocument36 pagesCell Specification MechanismsLily VaughNo ratings yet
- Vulval Development in C ElegansDocument28 pagesVulval Development in C ElegansSourav DeyNo ratings yet
- C.elegans model for developmentDocument6 pagesC.elegans model for developmentranjan sahuNo ratings yet
- Developmental Biology Test QuestionsDocument73 pagesDevelopmental Biology Test QuestionsPearl PascuaNo ratings yet
- Lo Sviluppo Della DrosophilaDocument32 pagesLo Sviluppo Della DrosophilaLorenzo MicheliNo ratings yet
- Animal Diversity and DevelopmentDocument68 pagesAnimal Diversity and DevelopmentGabrielle Salamanca CastuloNo ratings yet
- Methods of StudyDocument27 pagesMethods of StudyMary Eunice Elizabeth YraNo ratings yet
- Germ Cells 1Document5 pagesGerm Cells 1Attiya SabirNo ratings yet
- Germ cell migration and differentiation in amphibians, birds and mammalsDocument26 pagesGerm cell migration and differentiation in amphibians, birds and mammalsSales Him IT ServicesNo ratings yet
- LSM3233 Lecture 2 Notes 2013Document44 pagesLSM3233 Lecture 2 Notes 2013Prab NathanNo ratings yet
- Germline Regulatory Element of Oct-4 Specific For The Totipotent Cycle ofDocument14 pagesGermline Regulatory Element of Oct-4 Specific For The Totipotent Cycle ofMariane Gabriela Cesar Ribeiro FerreiraNo ratings yet
- Ans Key Mid Sem QP For DB 2019-20Document4 pagesAns Key Mid Sem QP For DB 2019-20Aaryan GuptaNo ratings yet
- Alan's Bio NotesDocument19 pagesAlan's Bio Notesluongc4100% (1)
- Key Events in DevelopmentDocument83 pagesKey Events in DevelopmentMelatiNo ratings yet
- Evaluation Component: Mid Semester (CB)Document5 pagesEvaluation Component: Mid Semester (CB)Aaryan GuptaNo ratings yet
- CAP 4 Development of The MammalianDocument12 pagesCAP 4 Development of The Mammaliangilson_pessoaNo ratings yet
- ADocument40 pagesAapi-3826751No ratings yet
- Pembelahan Embrio (Cleavage) S2 2017Document47 pagesPembelahan Embrio (Cleavage) S2 2017Desy NataliaNo ratings yet
- Caenorhabditis Elegans: Espineda. Estevanez. Natividad. UrbanoDocument20 pagesCaenorhabditis Elegans: Espineda. Estevanez. Natividad. UrbanoCourtneyNo ratings yet
- SummaryDocument5 pagesSummaryDuhreen Kate CastroNo ratings yet
- Role of Cytoplasm in Cell DifferentiationDocument7 pagesRole of Cytoplasm in Cell DifferentiationJ LALNo ratings yet
- 11 Axis FormationDocument61 pages11 Axis Formationapi-3800038100% (1)
- Moriyama 2018Document8 pagesMoriyama 2018Gabriel BuonoNo ratings yet
- BIO 325 Exam 1 ReviewDocument17 pagesBIO 325 Exam 1 ReviewNerdy Notes Inc.No ratings yet
- ANIMAL REPRODUCTIVE DEVELOPMENTDocument69 pagesANIMAL REPRODUCTIVE DEVELOPMENTRochelleCasador180No ratings yet
- GAMETOGENESISDocument44 pagesGAMETOGENESISMary Eunice Elizabeth YraNo ratings yet
- Molecular Basis of MorphogenesisDocument44 pagesMolecular Basis of MorphogenesisMitzel SapaloNo ratings yet
- Cell Differentiation Cmtut - 6305Document9 pagesCell Differentiation Cmtut - 6305rkblsistem100% (1)
- Topic 3 Revision: Structure of A Eukaryotic CellDocument10 pagesTopic 3 Revision: Structure of A Eukaryotic CellCam J BaileyNo ratings yet
- Early Development in InvertebratesDocument36 pagesEarly Development in Invertebratessiva100% (1)
- Developmental Biology H. 277 - Gillbert BarresiDocument57 pagesDevelopmental Biology H. 277 - Gillbert BarresiAndini M RNo ratings yet
- 5e's ActivitiesDocument2 pages5e's ActivitiesJoyae ChavezNo ratings yet
- Learning Models: Original Material by Dr. Mike Rapatan and Dr. Jade O. AlbertoDocument27 pagesLearning Models: Original Material by Dr. Mike Rapatan and Dr. Jade O. AlbertoJoyae ChavezNo ratings yet
- Learning Models: Original Material by Dr. Mike Rapatan and Dr. Jade O. AlbertoDocument27 pagesLearning Models: Original Material by Dr. Mike Rapatan and Dr. Jade O. AlbertoJoyae ChavezNo ratings yet
- Teaching Science Through Inquiry-1Document51 pagesTeaching Science Through Inquiry-1Joyae ChavezNo ratings yet
- Division Live - in Training & Writing On Interdisciplinary Contextualization & Inquiry - Based Science Teaching ApproachDocument8 pagesDivision Live - in Training & Writing On Interdisciplinary Contextualization & Inquiry - Based Science Teaching ApproachJoyae ChavezNo ratings yet
- 7 Es DLLDocument3 pages7 Es DLLJoyae ChavezNo ratings yet
- Inquiry-Based Teaching Plan-THIS Is FINAL NADocument13 pagesInquiry-Based Teaching Plan-THIS Is FINAL NAJoyae ChavezNo ratings yet
- Icon Ppt-This Is Final NaDocument27 pagesIcon Ppt-This Is Final NaJoyae ChavezNo ratings yet
- Inquiry PrimerDocument3 pagesInquiry PrimerJoyae ChavezNo ratings yet
- Plan developmentally sequenced teachingDocument32 pagesPlan developmentally sequenced teachingmaxy waveNo ratings yet
- Traaining Matrix IConDocument1 pageTraaining Matrix IConJoyae ChavezNo ratings yet
- Expanding The 5E ModelDocument5 pagesExpanding The 5E ModelJoyae ChavezNo ratings yet
- PPST - RP Module12, June2018 PDFDocument36 pagesPPST - RP Module12, June2018 PDFTeresa Medina Ticsay100% (3)
- Module9 PPST4 5 2Document36 pagesModule9 PPST4 5 2Jhonabie Suligan CadeliñaNo ratings yet
- Module11.PPST5 .2.2 PDFDocument36 pagesModule11.PPST5 .2.2 PDFGinny Paul Gabuan100% (1)
- Design, select, organize and use diagnostic, formative and summative assessment strategiesDocument36 pagesDesign, select, organize and use diagnostic, formative and summative assessment strategiesAlbert Quia100% (1)
- Module8 PPST4 4 2Document36 pagesModule8 PPST4 4 2Roseclyde Dela Cruz-LucasanNo ratings yet
- Engage learners through meaningful exploration and hands-on activitiesDocument36 pagesEngage learners through meaningful exploration and hands-on activitiesJhonabie Suligan Cadeliña100% (1)
- Module5 PPST2 6 2Document36 pagesModule5 PPST2 6 2Jhonabie Suligan Cadeliña100% (1)
- Module3 PPST1 5 2Document32 pagesModule3 PPST1 5 2Jhonabie Suligan Cadeliña100% (2)
- Engage learners through meaningful exploration and hands-on activitiesDocument36 pagesEngage learners through meaningful exploration and hands-on activitiesJhonabie Suligan Cadeliña100% (1)
- Differentiated learning experiencesDocument30 pagesDifferentiated learning experiencesRoel Etcobanez Versales75% (8)
- Enhance learner achievement in literacy and numeracyDocument32 pagesEnhance learner achievement in literacy and numeracyJhonabie Suligan Cadeliña100% (2)
- Enhance learner achievement in literacy and numeracyDocument32 pagesEnhance learner achievement in literacy and numeracyJhonabie Suligan Cadeliña100% (2)
- Module3 PPST1 5 2Document32 pagesModule3 PPST1 5 2Jhonabie Suligan Cadeliña100% (2)
- Module1 PPST1 1 2 PDFDocument36 pagesModule1 PPST1 1 2 PDFRoel Etcobanez Versales100% (3)
- Module1 PPST1 1 2 PDFDocument36 pagesModule1 PPST1 1 2 PDFRoel Etcobanez Versales100% (3)
- K To 12 CG Science Gr.7 v1.0Document19 pagesK To 12 CG Science Gr.7 v1.0leila DNo ratings yet
- Practical Test: Joy C. ChavezDocument2 pagesPractical Test: Joy C. ChavezJoyae ChavezNo ratings yet
- Improving Instructional Leadership Behaviors of School PrincipalsDocument130 pagesImproving Instructional Leadership Behaviors of School PrincipalsJoyae ChavezNo ratings yet
- 1 s2.0 S071734581630001X MainDocument7 pages1 s2.0 S071734581630001X MainTAUHID ALAMNo ratings yet
- Bio Practice TestDocument27 pagesBio Practice TestMichelleNo ratings yet
- Mathlearnermodule 130708064103 Phpapp02 PDFDocument112 pagesMathlearnermodule 130708064103 Phpapp02 PDFnel baradiNo ratings yet
- Beekeeping Bonga University, Ethiopia, Seid M.Document32 pagesBeekeeping Bonga University, Ethiopia, Seid M.seidNo ratings yet
- Sumayya Arif Roll# 19-11120 Organization:UVA S: BIOT 498 Internship Course Final PresentationDocument7 pagesSumayya Arif Roll# 19-11120 Organization:UVA S: BIOT 498 Internship Course Final PresentationSumayya ArifNo ratings yet
- Genes and Heredity: Lesson on Chromosomes, Genetic Variation, and MutationsTITLE Topic 4: Understanding How Genes and Chromosomes Influence Traits and HeredityDocument23 pagesGenes and Heredity: Lesson on Chromosomes, Genetic Variation, and MutationsTITLE Topic 4: Understanding How Genes and Chromosomes Influence Traits and HeredityMouza Mubarak Sultan Mohammed Al KhailiNo ratings yet
- Ethics and Cloning: Invited ReviewDocument7 pagesEthics and Cloning: Invited ReviewHammadNo ratings yet
- Histophysiology+of+the+Skin+for+Vula+ Dr+MowlaDocument28 pagesHistophysiology+of+the+Skin+for+Vula+ Dr+MowlaYaaseen GallantNo ratings yet
- ĐỀ SỐ 8Document5 pagesĐỀ SỐ 8Anh MaiNo ratings yet
- Pearson Edexcel International GCSE in Biology (9-1) Exemplar Student AnswersDocument59 pagesPearson Edexcel International GCSE in Biology (9-1) Exemplar Student AnswersPedroNo ratings yet
- BIOL 102 - Group Activity #2Document8 pagesBIOL 102 - Group Activity #2Abhi RajNo ratings yet
- AVB Sepharose High Performance: GE HealthcareDocument2 pagesAVB Sepharose High Performance: GE HealthcareMike BudimanNo ratings yet
- Genetic EngineeringDocument34 pagesGenetic EngineeringTiffany GordonNo ratings yet
- Biology 400 MCQs in English - (WWW - GkNotesPDF.com)Document26 pagesBiology 400 MCQs in English - (WWW - GkNotesPDF.com)jeetendra sahuNo ratings yet
- DuonsDocument7 pagesDuonsJulián Andrés Sánchez PachónNo ratings yet
- Paper 2 S W8Document13 pagesPaper 2 S W8Ahmed SahilNo ratings yet
- Lxad 105Document46 pagesLxad 105Samson Debbarma - M.Sc. MicrobiologyNo ratings yet
- Neuroscience and Biobehavioral ReviewsDocument34 pagesNeuroscience and Biobehavioral ReviewsRosa Indah KusumawardaniNo ratings yet
- Epidermal Permeability Barrier Defects and Barrier Repair Therapy in Atopic DermatitisDocument12 pagesEpidermal Permeability Barrier Defects and Barrier Repair Therapy in Atopic DermatitisMisaeldpdNo ratings yet
- Biology Mock Paper 3Document11 pagesBiology Mock Paper 3chemNo ratings yet
- Gene ExpressionDocument5 pagesGene ExpressionbenjaminlugahillaryNo ratings yet
- Finals Examination Biochemistry Lecture RNA Translation and MutationDocument7 pagesFinals Examination Biochemistry Lecture RNA Translation and MutationAlbertNo ratings yet
- CBSE Class 10 Science Sample Paper 5 SolutionsDocument4 pagesCBSE Class 10 Science Sample Paper 5 SolutionsSreehari.PNo ratings yet
- Student Exploration: Evolution-CladogramsDocument14 pagesStudent Exploration: Evolution-CladogramsAshNo ratings yet
- Visions of The Future WorksheetDocument2 pagesVisions of The Future WorksheetKevin Joseph PerfectoNo ratings yet
- Asexual Reproduction Worksheet Section A: Page No 147Document10 pagesAsexual Reproduction Worksheet Section A: Page No 147Christina T Z-chYnNo ratings yet
- Trends in Anaesthesia and Critical Care: Jamie Sleigh, Martyn Harvey, Logan Voss, Bill DennyDocument6 pagesTrends in Anaesthesia and Critical Care: Jamie Sleigh, Martyn Harvey, Logan Voss, Bill DennyAnonymous ORleRrNo ratings yet
- Molecular Biology Assignment QuestionsDocument26 pagesMolecular Biology Assignment QuestionsPARVATHY ANILNo ratings yet
- Bacterial InfectionsDocument6 pagesBacterial InfectionsErika Noreen Dela RosaNo ratings yet
- Kami Export - Luuk Van Meurs - DNA Replication WorksheetDocument2 pagesKami Export - Luuk Van Meurs - DNA Replication WorksheetLuuk Van MeursNo ratings yet