You might also like
- Cell PhysiologyDocument45 pagesCell PhysiologyHimani JhaNo ratings yet
- Basics of Human CellDocument24 pagesBasics of Human Cellananya girishbabuNo ratings yet
- 1.2 Cell PhysiologyDocument39 pages1.2 Cell PhysiologyChristone “Zuluzulu” ZuluNo ratings yet
- Hist of Cell-Organelles 2022Document45 pagesHist of Cell-Organelles 2022betelhema71No ratings yet
- Lecture Intro To Cell Physiology VersionDocument57 pagesLecture Intro To Cell Physiology Versionranart8100% (1)
- 4.cell 1Document67 pages4.cell 1Raymond LundaNo ratings yet
- Cell Components Structure and FunctionDocument45 pagesCell Components Structure and FunctionJada HartNo ratings yet
- 1 Lec.1 (w1) Overview and Plant StructureDocument32 pages1 Lec.1 (w1) Overview and Plant StructureRabiatul AdawiyyahNo ratings yet
- Biology ObjectivesDocument12 pagesBiology ObjectivesNicholas OwNo ratings yet
- Lecture 3 - Cell Structure & FunctionDocument27 pagesLecture 3 - Cell Structure & Functionlazysadoon09No ratings yet
- Cells: General ZoologyDocument71 pagesCells: General ZoologyAzaleaNo ratings yet
- Plant Cell Structure and FunctionDocument29 pagesPlant Cell Structure and Functionihebunnaogochukwu24No ratings yet
- Cell Structure and Function: Dr. N. OjehDocument45 pagesCell Structure and Function: Dr. N. OjehZachary HamidNo ratings yet
- Cell Structure & Function 2022Document43 pagesCell Structure & Function 2022Kavita MahaseNo ratings yet
- Cell Junction 2Document90 pagesCell Junction 2sohail shaikhNo ratings yet
- NUCLEUS NUCLEAR STRUCTURES and ORGANELLESDocument39 pagesNUCLEUS NUCLEAR STRUCTURES and ORGANELLESolawandeilo123No ratings yet
- S 13 Lect 2Document35 pagesS 13 Lect 2Anonymous fbXIfToSNo ratings yet
- Cell - The Unit of Life BST NotesDocument10 pagesCell - The Unit of Life BST NotesE.AshwinNo ratings yet
- General Medical Biology: Lecture 3: Cell Structure and FunctionDocument24 pagesGeneral Medical Biology: Lecture 3: Cell Structure and FunctionRasan QadrNo ratings yet
- Komar University Cell Structure and Function LectureDocument23 pagesKomar University Cell Structure and Function LectureRasan QadrNo ratings yet
- Cell Biology Lec 5Document18 pagesCell Biology Lec 5Mushtaq HassanNo ratings yet
- Chapter 2 Structure of Cells and OrganellesDocument42 pagesChapter 2 Structure of Cells and OrganellesAnonymous wj0YtncYYINo ratings yet
- Chapter 3-2Document62 pagesChapter 3-2Sifan MotumaNo ratings yet
- The Cell: and Cellular OrganellesDocument70 pagesThe Cell: and Cellular OrganellesKunda JosephNo ratings yet
- Cell PhysiologyDocument31 pagesCell PhysiologyJulie Calo PugiesNo ratings yet
- Human Cell AnatomyDocument56 pagesHuman Cell Anatomysanthiyasandy100% (3)
- Cell Structure and FunctionDocument32 pagesCell Structure and Functionnaveedalikhoso365100% (1)
- Chapter 3-Cell Structure and TaxonomyDocument60 pagesChapter 3-Cell Structure and TaxonomyMarcos AlbaridaNo ratings yet
- The Cell Structure, Functions of The CellDocument76 pagesThe Cell Structure, Functions of The CellCharles SsekawuNo ratings yet
- Cell Structure and TypeDocument9 pagesCell Structure and TypeFadhil Hussam AhmedNo ratings yet
- Cellular Level of OrganizationDocument72 pagesCellular Level of OrganizationPandit ManishNo ratings yet
- Prokaryotic and Eukaryotic CellsDocument78 pagesProkaryotic and Eukaryotic CellsLandon GlascockNo ratings yet
- Lesson 2 CELLDocument101 pagesLesson 2 CELLJulius Memeg PanayoNo ratings yet
- Cell 2Document45 pagesCell 2Majd HusseinNo ratings yet
- Pathophysiology: General Concept of Diseases in CellDocument72 pagesPathophysiology: General Concept of Diseases in Cellbrahmani ptrNo ratings yet
- Cytoplasm By: Prof. Dr. Suzi H. Ibrahim Lectures 1,2&3Document58 pagesCytoplasm By: Prof. Dr. Suzi H. Ibrahim Lectures 1,2&3eman el saeedNo ratings yet
- Cellular Structure: A Guide to the Organelles and Components of the CellDocument24 pagesCellular Structure: A Guide to the Organelles and Components of the CellTyra Joyce Revadavia100% (1)
- Endomembrane System (Seminar)Document11 pagesEndomembrane System (Seminar)Prachi TayadeNo ratings yet
- K.2-4 Organisasi SelDocument82 pagesK.2-4 Organisasi SelSamuel Assan SiraitNo ratings yet
- Pengantar Biologi SelDocument81 pagesPengantar Biologi SeldiesNo ratings yet
- 4, 5 - The Cytoplasm - 2Document58 pages4, 5 - The Cytoplasm - 2Muthana Bani YassenNo ratings yet
- Structure of CellDocument42 pagesStructure of CellJeevitha VanithaNo ratings yet
- CellDocument5 pagesCellhkrybmzxfbxbwnpfhnNo ratings yet
- Cell MembraneDocument100 pagesCell MembranephyMedza AppNo ratings yet
- General Physiology: First YearDocument67 pagesGeneral Physiology: First YearGo HellNo ratings yet
- Cytoplasmic Membrane SystemDocument58 pagesCytoplasmic Membrane Systemsjbiotech0% (1)
- Cell Structure: A Guide to the OrganellesDocument11 pagesCell Structure: A Guide to the OrganellesAli MahmoudNo ratings yet
- Membrane PhysiologyDocument223 pagesMembrane PhysiologyteklayNo ratings yet
- Structure and Function of Cell: Presented by Miss - Robinsini.F 4 Year BSC Nursing Hosmat College of NursingDocument29 pagesStructure and Function of Cell: Presented by Miss - Robinsini.F 4 Year BSC Nursing Hosmat College of NursingJoshua SpotifyNo ratings yet
- Notes CH 1 and 2Document59 pagesNotes CH 1 and 2Salman AbidNo ratings yet
- Lecture 2 - Cell PhysiologyDocument12 pagesLecture 2 - Cell Physiologyyaqooburooj18No ratings yet
- Cells: Cytoplasmic OrganellesDocument7 pagesCells: Cytoplasmic OrganellesJonathan HigginbothamNo ratings yet
- Cell Structure 3Document30 pagesCell Structure 3Rohan PaneruNo ratings yet
- Cell HistologyDocument16 pagesCell Histologypauline_balidiongNo ratings yet
- Cell & Molecular PhysiologyDocument129 pagesCell & Molecular PhysiologyDr Imran AftabNo ratings yet
- Physio 2 Cell PhysiologyDocument60 pagesPhysio 2 Cell PhysiologyAnonymous t5TDwdNo ratings yet
- Screenshot 2021-02-08 at 9.24.46 PMDocument21 pagesScreenshot 2021-02-08 at 9.24.46 PMsarwaNo ratings yet
- Review of Cell Biology Concepts: Differences Between Prokaryotic Vs Eukaryotic CellsDocument25 pagesReview of Cell Biology Concepts: Differences Between Prokaryotic Vs Eukaryotic Cellsevil twinNo ratings yet
- General Physiology by BrownDocument51 pagesGeneral Physiology by BrownuzohchukwuebukaNo ratings yet
- Development and Course: TemperamentalDocument4 pagesDevelopment and Course: TemperamentalAuliya SyismaNo ratings yet
- Surviving Sepsis Campaign International.21Document81 pagesSurviving Sepsis Campaign International.21jamesteryNo ratings yet
- Interaksi Antar Sel Dan Transduksi SinyalDocument55 pagesInteraksi Antar Sel Dan Transduksi SinyalRachmaningtyas Setyo Hartono PutriNo ratings yet
- Family Medicine: Nendyah RoestijawatiDocument17 pagesFamily Medicine: Nendyah RoestijawatiAuliya SyismaNo ratings yet
- PCOSDocument1 pagePCOSAuliya SyismaNo ratings yet
- Daftar PustakaDocument1 pageDaftar PustakaAuliya SyismaNo ratings yet
- Ultrasound-Guided Interventional Procedures in Pain ManagementDocument4 pagesUltrasound-Guided Interventional Procedures in Pain ManagementAuliya SyismaNo ratings yet
- Manage Pain in LTCDocument40 pagesManage Pain in LTCPutra Pratama100% (1)
- Jenis Kelamin, Usia, Pengetahuan, Ekonomi, Pola Makan, dan Makanan Iritatif terhadap Status GastritisDocument16 pagesJenis Kelamin, Usia, Pengetahuan, Ekonomi, Pola Makan, dan Makanan Iritatif terhadap Status GastritisRio Taruna JatiNo ratings yet
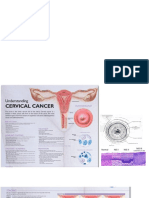
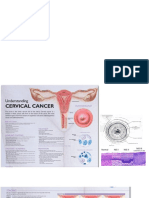
- Ca CerviksDocument5 pagesCa CerviksAuliya SyismaNo ratings yet
- CA CerviksDocument5 pagesCA CerviksAuliya SyismaNo ratings yet
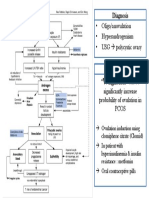
- Weight Loss: Diagnosis - Oligo/anovulation - Hyperandrogenism - USG Polycystic OvaryDocument1 pageWeight Loss: Diagnosis - Oligo/anovulation - Hyperandrogenism - USG Polycystic OvaryAuliya SyismaNo ratings yet
- Eczema HerpeticumDocument4 pagesEczema HerpeticumAuliya SyismaNo ratings yet
- Max Gerson - The Gerson Therapy Has Proved EffectiveDocument8 pagesMax Gerson - The Gerson Therapy Has Proved EffectiveFernando CordeiroNo ratings yet
- The Ultimate A-Level Aqa Biology Cheatsheet PackDocument17 pagesThe Ultimate A-Level Aqa Biology Cheatsheet PackJamesNo ratings yet
- Lab Worksheet No. 3 The CellDocument20 pagesLab Worksheet No. 3 The CellKarylle Ezra GulifardoNo ratings yet
- Activation of Our Crystalline Sun DNA TemplatesDocument28 pagesActivation of Our Crystalline Sun DNA TemplatesfiremanM100% (16)
- All Questions of Life Science III +2021Document147 pagesAll Questions of Life Science III +2021lashqarghanaiymNo ratings yet
- BILD 3 Practice Final FA08Document10 pagesBILD 3 Practice Final FA08George Chen100% (1)
- The Complete Biomonster Vocabulary Reference GuideDocument26 pagesThe Complete Biomonster Vocabulary Reference Guideapi-110789702100% (1)
- Animal and Plant CellDocument41 pagesAnimal and Plant CellSabrina LavegaNo ratings yet
- PT1 Data Sheet Cell Structure - Examining Plant and Animal CellsDocument8 pagesPT1 Data Sheet Cell Structure - Examining Plant and Animal CellsAsuka ChanNo ratings yet
- Dr. Retno Sutomo, PH.D, Sp.ADocument37 pagesDr. Retno Sutomo, PH.D, Sp.ADoni RevaiNo ratings yet
- PHTH 211 Cell Membrane PhysiologyDocument36 pagesPHTH 211 Cell Membrane Physiologymaatela7sasNo ratings yet
- Lakhmir Singh Biology Class 9 Solutions CH 2Document7 pagesLakhmir Singh Biology Class 9 Solutions CH 2DarshilNo ratings yet
- Moleculer BiologyDocument159 pagesMoleculer BiologyVeekeshGupta100% (3)
- Introduction To Cells PDFDocument6 pagesIntroduction To Cells PDFZainab HamidNo ratings yet
- A Cell Is Like A FactoryDocument13 pagesA Cell Is Like A FactoryNoralyn Ngislawan-GunnawaNo ratings yet
- SEMINAR REPORT TODAY 3 Almost Done 1Document21 pagesSEMINAR REPORT TODAY 3 Almost Done 1Afsal ShajiNo ratings yet
- Unit 2 - Introduction To BiochemistryDocument8 pagesUnit 2 - Introduction To BiochemistryWinrich Louise M. MontanoNo ratings yet
- B.sc. Hons. ZoologyDocument110 pagesB.sc. Hons. ZoologyHarsh Dalal0% (1)
- Read It! Specialised CellsDocument6 pagesRead It! Specialised CellsVictoria LowmanNo ratings yet
- Fifth Edition Answers To End-Of-Chapter Questions: GCSE Biology For YouDocument1 pageFifth Edition Answers To End-Of-Chapter Questions: GCSE Biology For YouclydeNo ratings yet
- MCQs About Cell BiologyDocument3 pagesMCQs About Cell BiologyHeba M.abueyadaNo ratings yet
- The Main Themes of MicrobiologyDocument34 pagesThe Main Themes of MicrobiologyNurseReyesNo ratings yet
- Cells: A Basic IntroductionDocument18 pagesCells: A Basic IntroductionMuqadasNo ratings yet
- MSc Biotechnology Scheme of Studies & Exams 2012-13Document39 pagesMSc Biotechnology Scheme of Studies & Exams 2012-13DeepakNo ratings yet
- Test Bank For Davis Advantage For Pathophysiology: Introductory Concepts and Clinical Perspectives, 2nd Edition, Theresa CapriottiDocument23 pagesTest Bank For Davis Advantage For Pathophysiology: Introductory Concepts and Clinical Perspectives, 2nd Edition, Theresa Capriottinombril.skelp15v4100% (15)
- Essential, Trace, and Nonessential Ions-1Document3 pagesEssential, Trace, and Nonessential Ions-1John AndanNo ratings yet
- For Biology 250 at CSM: Compiled From Marieb's Human Anatomy & PhysiologyDocument26 pagesFor Biology 250 at CSM: Compiled From Marieb's Human Anatomy & PhysiologycathjoyceNo ratings yet
- Diffusion Pre LabDocument2 pagesDiffusion Pre Labkina lovleyNo ratings yet
- Nhóm 6 - Fucoidan and Treatment CancerDocument18 pagesNhóm 6 - Fucoidan and Treatment CancerThị ThiênNo ratings yet
- Dwnload Full Psychology 8th Edition Gleitman Test Bank PDFDocument35 pagesDwnload Full Psychology 8th Edition Gleitman Test Bank PDFiseasfoqarae100% (10)