You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5819)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (845)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- MCQs On Circulation Physiology With KeyDocument6 pagesMCQs On Circulation Physiology With KeyMudassar Roomi92% (36)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sea Urchin Fertilization LabDocument5 pagesThe Sea Urchin Fertilization LabAmy HollingsworthNo ratings yet
- Pentagon Maternal and Child PDFDocument38 pagesPentagon Maternal and Child PDFmarie89meNo ratings yet
- Histology: Lecture NotesDocument97 pagesHistology: Lecture Notessky100% (4)
- Slice of LifeDocument27 pagesSlice of Lifejhjg9089No ratings yet
- ApoptosisDocument23 pagesApoptosisLavanya Kalapala100% (1)
- Rishikesh Seminar Content - SSSOHADocument13 pagesRishikesh Seminar Content - SSSOHARabindra DhitalNo ratings yet
- Sensual Massage For Couples (PDFDrive)Document196 pagesSensual Massage For Couples (PDFDrive)Lauren Xavier100% (3)
- TympanoplastyDocument75 pagesTympanoplastylisabdaNo ratings yet
- Hipertiroidismo y EmbarazoDocument12 pagesHipertiroidismo y EmbarazoAlexy SanchezNo ratings yet
- Rajah 1.1 Menunjukkan Tindakan Enzim Maltase Ke Atas Substrat PDocument3 pagesRajah 1.1 Menunjukkan Tindakan Enzim Maltase Ke Atas Substrat PAZIANA YUSUFNo ratings yet
- An Assignment On Autoimmunity: July 2012Document15 pagesAn Assignment On Autoimmunity: July 2012SudhakarNo ratings yet
- Structural Organisation in Animals: Morphology and Anatomy of CockroachDocument13 pagesStructural Organisation in Animals: Morphology and Anatomy of CockroachKali Shankar SinghNo ratings yet
- Ear Histology: Ratnafitrir. Histology Laboratory UnissulaDocument53 pagesEar Histology: Ratnafitrir. Histology Laboratory UnissulaRatna Fitri RahayuningsihNo ratings yet
- Kingdom Animalia NotesDocument2 pagesKingdom Animalia Notesjoann thomasNo ratings yet
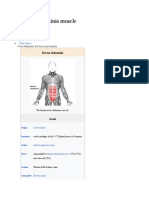
- Rectus Abdominis MuscleDocument3 pagesRectus Abdominis MuscleBatang HamogNo ratings yet
- Feline Orthopaedics, 2nd Edition (VetBooks - Ir)Document400 pagesFeline Orthopaedics, 2nd Edition (VetBooks - Ir)Narvarte Hospital Veterinario de EspecialidadesNo ratings yet
- (IMMUNOHEMA) Denise M. Harmening - Modern Blood Banking & Transfusion Practices (2019, F.A. Davis)Document56 pages(IMMUNOHEMA) Denise M. Harmening - Modern Blood Banking & Transfusion Practices (2019, F.A. Davis)Who KnowsNo ratings yet
- Sexual Reproduction in Flowering PlantsDocument35 pagesSexual Reproduction in Flowering PlantsDiksha TNo ratings yet
- The Allergy Book - Solving Your Family's Nasal Allergies, Asthma, Food Sensitivities, and Related Health and Behavioral Problems (PDFDrive)Document282 pagesThe Allergy Book - Solving Your Family's Nasal Allergies, Asthma, Food Sensitivities, and Related Health and Behavioral Problems (PDFDrive)angelobuffaloNo ratings yet
- Transport in Animals 1 StructuredDocument12 pagesTransport in Animals 1 Structuredlogy of bioNo ratings yet
- Bio ACE Form 4Document3 pagesBio ACE Form 4Myramel KlarisNo ratings yet
- Fetal Microchimerism and Maternal HealthDocument13 pagesFetal Microchimerism and Maternal HealthFranco Fabricio CarpioNo ratings yet
- Interpretation of Cervical X-RayDocument31 pagesInterpretation of Cervical X-RayblapuNo ratings yet
- Tissue Matching Worksheet: 1. Match Each General Tissue Category To The Appropriate FeatureDocument5 pagesTissue Matching Worksheet: 1. Match Each General Tissue Category To The Appropriate FeatureAnghel LopezNo ratings yet
- Actividad Semana 1 - Anatomia y Fisiologia.Document7 pagesActividad Semana 1 - Anatomia y Fisiologia.Claribel Sarait Blanco GiilNo ratings yet
- Immunoglobulins - Structure and Function Definition: Immunoglobulins (Ig)Document9 pagesImmunoglobulins - Structure and Function Definition: Immunoglobulins (Ig)Valdez Francis ZaccheauNo ratings yet
- Muscle ContractionDocument2 pagesMuscle Contractionaikoii sanNo ratings yet
- Coagulation DisordersDocument26 pagesCoagulation DisordersLia pramita0% (1)
- Anatomy of AirwaysDocument6 pagesAnatomy of Airwaysgdubs215No ratings yet