You might also like
- Character and Plot Lesson PlanDocument7 pagesCharacter and Plot Lesson Planapi-274109706No ratings yet
- A New General Catalogue of The Ants of The WorldDocument513 pagesA New General Catalogue of The Ants of The WorldFabiana Soares Cariri Lopes100% (1)
- 04-Coats and Clamp 2009-Ciliated Protists of The GoMxDocument24 pages04-Coats and Clamp 2009-Ciliated Protists of The GoMxebutaNo ratings yet
- Good Songs For Early YearsDocument18 pagesGood Songs For Early YearsjackmcfrenzieNo ratings yet
- MTG CocktailsDocument21 pagesMTG CocktailsGianniNo ratings yet
- Marshal and Thornton 1963. MICROMALTHUS (Coleoptera - Micromalthidae) in Hong KongDocument6 pagesMarshal and Thornton 1963. MICROMALTHUS (Coleoptera - Micromalthidae) in Hong KongCarlos AguilarNo ratings yet
- Crustacea: Non-Cladoceran Branchiopoda: Joel W. Martin and Sarah L. BoyceDocument14 pagesCrustacea: Non-Cladoceran Branchiopoda: Joel W. Martin and Sarah L. BoyceDhimas AfihandarinNo ratings yet
- Phylum ArthropodaDocument19 pagesPhylum ArthropodaOkta Novalia GNo ratings yet
- EdeagosDocument8 pagesEdeagosAdriana EcheverríaNo ratings yet
- Fryda&Rohr2006shell HeterostrophyordovicianDocument8 pagesFryda&Rohr2006shell HeterostrophyordovicianGil Iack XimenesNo ratings yet
- ZootaxaDocument17 pagesZootaxaoliviamuliawatideviNo ratings yet
- 4a Polychaetes 03 Pogonophora 02Document30 pages4a Polychaetes 03 Pogonophora 02Wilson David Martínez HaadNo ratings yet
- Morph Amph Larv Dodd BookDocument16 pagesMorph Amph Larv Dodd BookMorgana OliveiraNo ratings yet
- Myxine Clave Affinis Australis Hubbsi HubbsoidesDocument21 pagesMyxine Clave Affinis Australis Hubbsi Hubbsoidesvictor_pasten_m6679No ratings yet
- Entomology 322 Lab 2 Introduction To Arthropoda: HexapodaDocument10 pagesEntomology 322 Lab 2 Introduction To Arthropoda: HexapodaHumberto GiraldoNo ratings yet
- Helicoplacus 2022Document15 pagesHelicoplacus 2022Amanda AngelesNo ratings yet
- MouthpartsDocument14 pagesMouthpartsjhosepNo ratings yet
- Braconidae1993ill Key PDFDocument187 pagesBraconidae1993ill Key PDFNicolas David Gutierrez RubianoNo ratings yet
- Brach IopodsDocument13 pagesBrach IopodsAthaariq AriqNo ratings yet
- New World AgaoninaeDocument53 pagesNew World Agaoninaeapi-3726790No ratings yet
- Morphology, Systematics, and Biology of The Spanish MackerelsDocument148 pagesMorphology, Systematics, and Biology of The Spanish MackerelsRio AlmusataNo ratings yet
- PlanktonDocument12 pagesPlanktonRen ObeñaNo ratings yet
- Australian Dragonflies: A Guide to the Identification, Distributions and Habitats of Australian OdonataFrom EverandAustralian Dragonflies: A Guide to the Identification, Distributions and Habitats of Australian OdonataRating: 3 out of 5 stars3/5 (1)
- Snakeheads ChannidaeDocument47 pagesSnakeheads ChannidaeAbdul HameedNo ratings yet
- Trichoptera Phylogeny Adaptations and BiogeographyDocument13 pagesTrichoptera Phylogeny Adaptations and BiogeographyCarli RodríguezNo ratings yet
- Trichoptera Phylogeny Adaptations and BiogeographyDocument13 pagesTrichoptera Phylogeny Adaptations and BiogeographyCarli RodríguezNo ratings yet
- The Origin and Evolution of The Angiosperm Carpel: David Winship Taylor and Gretchen KirchnerDocument25 pagesThe Origin and Evolution of The Angiosperm Carpel: David Winship Taylor and Gretchen KirchnerDiana MendoncaNo ratings yet
- Tunicata: Alberto Stolfi and Federico D. BrownDocument70 pagesTunicata: Alberto Stolfi and Federico D. BrownAmalia Nur LatifahNo ratings yet
- 1988 ClarkDocument44 pages1988 ClarkDiego MancillaNo ratings yet
- Full Download Book Linear Algebra and Its Applications Global Edition PDFDocument22 pagesFull Download Book Linear Algebra and Its Applications Global Edition PDFheather.hermosillo571100% (20)
- The Theropod FuculaDocument24 pagesThe Theropod FuculaMyrthe de GoeijNo ratings yet
- Boldrini Et Al., 2017 (Brasil) BaDocument10 pagesBoldrini Et Al., 2017 (Brasil) Bacarmen villalobosNo ratings yet
- Pleistocene Pocket Gophers From San Josecito Cave, Nuevo Leon, MexicoFrom EverandPleistocene Pocket Gophers From San Josecito Cave, Nuevo Leon, MexicoNo ratings yet
- Knight (Kunci SergestidaeDocument27 pagesKnight (Kunci SergestidaeHarditya FirdhausNo ratings yet
- Crustaceans: Nicolas RabetDocument17 pagesCrustaceans: Nicolas RabetSarath KumaraNo ratings yet
- Artificial Keys To The Genera of Living Stalked CRDocument33 pagesArtificial Keys To The Genera of Living Stalked CRAurelia Izzatul AzkaNo ratings yet
- Laventa TitlepageDocument4 pagesLaventa TitlepageTefaNo ratings yet
- The Lagenar Otoliths of Teleosts: Their Morphology and Its Application in Species Identification, Phylogeny and SystematicsDocument28 pagesThe Lagenar Otoliths of Teleosts: Their Morphology and Its Application in Species Identification, Phylogeny and SystematicsMoisés Aldana AcostaNo ratings yet
- The Ecological Role of The Callitrichidae A Review (1984) R.W. SUSSMAN AND W.G. KINZEY.Document31 pagesThe Ecological Role of The Callitrichidae A Review (1984) R.W. SUSSMAN AND W.G. KINZEY.Sandy JP Souza100% (1)
- Nelson - 1985 - Some Characters of TrichonotidaeDocument6 pagesNelson - 1985 - Some Characters of TrichonotidaeMuntasir AkashNo ratings yet
- 026 Gullan Kondo 2008 Lac Insect MorphologyDocument8 pages026 Gullan Kondo 2008 Lac Insect MorphologySaneela NotaniNo ratings yet
- Fauna Amphibia GeneralDocument14 pagesFauna Amphibia GeneralCamilleLouiseJaureguiNo ratings yet
- Non Mammalian BarbelsDocument41 pagesNon Mammalian BarbelsDamian HaydenNo ratings yet
- Morphology and Molecular Phylogeny of Peritrich Ciliate Epibionts On Pelagic Diatoms: Vorticella Oceanica and Pseudovorticella Coscinodisci Sp. Nov. (Ciliophora, Peritrichia)Document12 pagesMorphology and Molecular Phylogeny of Peritrich Ciliate Epibionts On Pelagic Diatoms: Vorticella Oceanica and Pseudovorticella Coscinodisci Sp. Nov. (Ciliophora, Peritrichia)fernando_gomez8953No ratings yet
- Beetles Insecta ColeopteraDocument14 pagesBeetles Insecta Coleopteradipeshsingh563No ratings yet
- AnfibiosDocument31 pagesAnfibiosMary M. Gonzalez100% (1)
- A New Mourasuchus (Alligatoroidea, Caimaninae) From The Late Miocene of Venezuela, The Phylogeny of Caimaninae and Considerations On The Feeding Habits of Mourasuchus - PMCDocument41 pagesA New Mourasuchus (Alligatoroidea, Caimaninae) From The Late Miocene of Venezuela, The Phylogeny of Caimaninae and Considerations On The Feeding Habits of Mourasuchus - PMCHyperionEremitaNo ratings yet
- Morfologi (Sertakan Gambar Beserta Rincian Bagian-Bagian Tubuh Yang Di Riview) Dan Fungsi Bagian TubuhDocument13 pagesMorfologi (Sertakan Gambar Beserta Rincian Bagian-Bagian Tubuh Yang Di Riview) Dan Fungsi Bagian TubuhAMELIA ULMNo ratings yet
- Dinosaur PsittacosaurusDocument5 pagesDinosaur PsittacosaurusKevin Diego Hernandez MenaNo ratings yet
- Further Twists in Gastropod Shell EvolutionDocument4 pagesFurther Twists in Gastropod Shell EvolutionLeoregine RodriguezNo ratings yet
- The Taxonomic Position: Phaeodactylum TricornutumDocument7 pagesThe Taxonomic Position: Phaeodactylum TricornutumPatrícia BentoNo ratings yet
- Frey & Salisbury - 2007 - Chapter 16 - in Martill Et Al (Crocs Crato FM)Document16 pagesFrey & Salisbury - 2007 - Chapter 16 - in Martill Et Al (Crocs Crato FM)Crocodilo VasconcelosNo ratings yet
- Stone FliesDocument23 pagesStone Flieslottie.100% (1)
- Phylogeny of The Terrestrial IsopodaDocument36 pagesPhylogeny of The Terrestrial IsopodaPedroNo ratings yet
- Rotifera From Australian Inland Waters IDocument30 pagesRotifera From Australian Inland Waters ImeryNo ratings yet
- The British Woodlice: Being a Monograph of the Terrestrial Isopod Crustacea Occurring in the British IslandsFrom EverandThe British Woodlice: Being a Monograph of the Terrestrial Isopod Crustacea Occurring in the British IslandsNo ratings yet
- Phyletic Relationships of Living Sharks and RaysDocument20 pagesPhyletic Relationships of Living Sharks and RaysZAk1No ratings yet
- Thoracic and Coracoid Arteries In Two Families of Birds, Columbidae and HirundinidaeFrom EverandThoracic and Coracoid Arteries In Two Families of Birds, Columbidae and HirundinidaeNo ratings yet
- Laventa TitlepageDocument4 pagesLaventa TitlepageAlejandra SandovalNo ratings yet
- Economic Potential of Marble From Obajana, North Central, NigeriaDocument2 pagesEconomic Potential of Marble From Obajana, North Central, NigeriaAmri RizalNo ratings yet
- Fsdklfdofjew Lgfqeklfghe Wifss Skfn3ekfvcw Kfievkfiheifk .Dekegnow4 RSDNG 43rdDocument2 pagesFsdklfdofjew Lgfqeklfghe Wifss Skfn3ekfvcw Kfievkfiheifk .Dekegnow4 RSDNG 43rdAmri RizalNo ratings yet
- Peta KavlingDocument1 pagePeta KavlingAmri RizalNo ratings yet
- MAPSDocument1 pageMAPSAmri RizalNo ratings yet
- Zjsxaliavnm C Lfseawefv Wnggfmcse Wajwefhevc Efngirgkngn VRVMCDKFNCR CgegegegircDocument2 pagesZjsxaliavnm C Lfseawefv Wnggfmcse Wajwefhevc Efngirgkngn VRVMCDKFNCR CgegegegircAmri RizalNo ratings yet
- Laboratorium Geologi Optik: NIM: 13/346803/TK/40648) Thin Sec. Code: ODocument2 pagesLaboratorium Geologi Optik: NIM: 13/346803/TK/40648) Thin Sec. Code: OAmri RizalNo ratings yet
- Published By: On Jun 02, 2012: NabellanurulfitriDocument2 pagesPublished By: On Jun 02, 2012: NabellanurulfitriAmri RizalNo ratings yet
- The Croods - Lessons (Final)Document54 pagesThe Croods - Lessons (Final)Khun Win MaungNo ratings yet
- The Three Fat MenDocument84 pagesThe Three Fat MenDan Vie100% (1)
- Korte 2005Document36 pagesKorte 2005CellaNo ratings yet
- BACTERIADocument13 pagesBACTERIAJohn Bradley GomezNo ratings yet
- Year2 PSY286 Observing SniffyDocument20 pagesYear2 PSY286 Observing Sniffyalancummins3556100% (1)
- HITUNG JUMLAH RBCDocument26 pagesHITUNG JUMLAH RBCLina NurkhasanahNo ratings yet
- Lotr - Extras - Battle Companies - Istari CreatorDocument11 pagesLotr - Extras - Battle Companies - Istari CreatorRon Van 't VeerNo ratings yet
- Meredith Et Al. 2011. Mammal Family Level Phylogeny. ScienceDocument5 pagesMeredith Et Al. 2011. Mammal Family Level Phylogeny. ScienceLinethVelasquezNo ratings yet
- A New Discovery of A Miracle in The QuranDocument7 pagesA New Discovery of A Miracle in The QuranalkeilaniNo ratings yet
- Soal Pts Big Kelas X Aph 22-1Document7 pagesSoal Pts Big Kelas X Aph 22-1Rizqy Achmad FattahNo ratings yet
- Teaching Plan On ConstipationDocument4 pagesTeaching Plan On ConstipationShumaila Alvi0% (1)
- An Overview of Oral TissuesDocument4 pagesAn Overview of Oral TissuesRenukaChauveNo ratings yet
- 1 - Gulfstream Park DRFDocument3 pages1 - Gulfstream Park DRFTortolero CesarNo ratings yet
- Calcium Levels and Physical Characteristics of Egg Poultry Shell With Soaking Various SolventDocument12 pagesCalcium Levels and Physical Characteristics of Egg Poultry Shell With Soaking Various SolventRizky BagasNo ratings yet
- Prevalence of Toxoplasma Gondii Infection in Domestic Animals in District Bannu Khyber Pakhtunkhwa (KP), PakistanDocument6 pagesPrevalence of Toxoplasma Gondii Infection in Domestic Animals in District Bannu Khyber Pakhtunkhwa (KP), PakistanOpenaccess Research paperNo ratings yet
- Laeral Silverhand - S Notebook - Surviving Icewind DaleDocument54 pagesLaeral Silverhand - S Notebook - Surviving Icewind DaleUranuz100% (1)
- Lab 11 PlatyhelminthesDocument20 pagesLab 11 PlatyhelminthesdraykidNo ratings yet
- Earth Day LiturgyDocument4 pagesEarth Day LiturgyInnu TuduNo ratings yet
- Loch Melfort HotelDocument1 pageLoch Melfort HotelHotelPRNo ratings yet
- IGCSE Edexcel 4 1 ExcretionDocument30 pagesIGCSE Edexcel 4 1 ExcretionRick WuNo ratings yet
- May-June 2009 Passages Newsletter, Pennsylvania Association For Sustainable AgricultureDocument32 pagesMay-June 2009 Passages Newsletter, Pennsylvania Association For Sustainable AgriculturePennsylvania Association for Sustainable AgricultureNo ratings yet
- White TigersDocument71 pagesWhite TigersWishal Shahi100% (1)
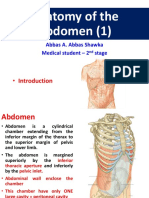
- Anatomy of The Abdomen (1) : - IntroductionDocument21 pagesAnatomy of The Abdomen (1) : - IntroductionMunachande KanondoNo ratings yet
- Microsurgical Anatomic Features of The Lamina TerminalisDocument8 pagesMicrosurgical Anatomic Features of The Lamina Terminalisghostmmx100% (2)
- REPORT TEXT AssignmentDocument5 pagesREPORT TEXT AssignmentDwi AgusNo ratings yet
- Portfolio Sleep DiaryDocument5 pagesPortfolio Sleep Diaryapi-301810518No ratings yet
- US Nursery Pigs M. Hyo PRDC Case Study (Roberts, 2011)Document4 pagesUS Nursery Pigs M. Hyo PRDC Case Study (Roberts, 2011)nick224No ratings yet