You might also like
- GED Science Practice Test Question Answers: Which Two Liquids Could Undergo An Acid Base Reaction?Document9 pagesGED Science Practice Test Question Answers: Which Two Liquids Could Undergo An Acid Base Reaction?Keny100% (1)
- Animal Biology Textbook Covers Key ConceptsDocument433 pagesAnimal Biology Textbook Covers Key Conceptsumairpk2011No ratings yet
- Fertility HerbsDocument17 pagesFertility HerbsKyle J. Norton100% (1)
- Isolation of Genomic DNA from Animal TissueDocument6 pagesIsolation of Genomic DNA from Animal TissueSanmugam SubbaramaniamNo ratings yet
- Regulation of the Cell Cycle by Protein KinasesDocument5 pagesRegulation of the Cell Cycle by Protein KinasesmohsinNo ratings yet
- Types of MutationsDocument25 pagesTypes of Mutationsrohishaak100% (1)
- Recombinant Protein Production in E.coliDocument23 pagesRecombinant Protein Production in E.coliSanjay KumarNo ratings yet
- Cloning Dolly & MicromanipulationDocument29 pagesCloning Dolly & Micromanipulationnitinyadav16No ratings yet
- Cell Cycle Cell Division PDFDocument13 pagesCell Cycle Cell Division PDFmuzamil shabir100% (1)
- Semen Analysis: Verian John Hoo - Sudario, RMTDocument37 pagesSemen Analysis: Verian John Hoo - Sudario, RMTepson printer100% (1)
- Overview of Cell Cycle by Javali.GDocument15 pagesOverview of Cell Cycle by Javali.GJavali.GNo ratings yet
- Protoplast Preparation and Somatic Hybrids: by Prof. M. Z. AbdinDocument49 pagesProtoplast Preparation and Somatic Hybrids: by Prof. M. Z. AbdinOriginate RecordsNo ratings yet
- lectut-BTN-303-pdf-Protoplast Technology PDFDocument12 pageslectut-BTN-303-pdf-Protoplast Technology PDFqwertNo ratings yet
- Basics of Cell Culture in 40 CharactersDocument33 pagesBasics of Cell Culture in 40 CharactersSai SridharNo ratings yet
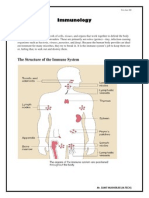
- Immunology NotesDocument8 pagesImmunology NotesSumit Mukherjee100% (1)
- Module-3 (Theory) Upstream and Down Stream Component of A Fermentation Process PDFDocument6 pagesModule-3 (Theory) Upstream and Down Stream Component of A Fermentation Process PDFAnonymous AgGBWdNo ratings yet
- Introduction to Biology in 40 CharactersDocument46 pagesIntroduction to Biology in 40 CharactersLawrence AnDrew FrondaNo ratings yet
- CryopreservationDocument29 pagesCryopreservationDharmendra KumarNo ratings yet
- Recombinant Protein ProductionDocument23 pagesRecombinant Protein ProductionAnamitra SenNo ratings yet
- ElisaDocument30 pagesElisaasal896059No ratings yet
- Genetics, Lecture 11 (Lecture Notes)Document16 pagesGenetics, Lecture 11 (Lecture Notes)Ali Al-QudsiNo ratings yet
- Plant MetabolitesDocument47 pagesPlant MetabolitesSrinivas Rallapalli100% (1)
- Composition of Milk PDFDocument5 pagesComposition of Milk PDFghanashyam tomarNo ratings yet
- Cellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsFrom EverandCellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsCharles G. CochraneNo ratings yet
- Gene Cloning TechnologyDocument16 pagesGene Cloning TechnologyNguyen Thanh Anh NiiroNo ratings yet
- Piezo-Actuated Mouse Intracytoplasmic Sperm Injection (ICSI)Document9 pagesPiezo-Actuated Mouse Intracytoplasmic Sperm Injection (ICSI)Juan De Dios HourcadeNo ratings yet
- 2t ScienceDocument96 pages2t SciencetoñiNo ratings yet
- Dynamics of The Mammalian Sperm Plasma Membrane in The Process of FertilizationDocument39 pagesDynamics of The Mammalian Sperm Plasma Membrane in The Process of FertilizationWachiel ArhamzNo ratings yet
- Fundamental GeneticsDocument14 pagesFundamental GeneticsjuvemaNo ratings yet
- VNS Faculty of Pharmacy: Mentors - Presented byDocument1 pageVNS Faculty of Pharmacy: Mentors - Presented bypoplu100% (1)
- Animal Cell CultureDocument33 pagesAnimal Cell CultureMd. Babul AktarNo ratings yet
- Organic AcidDocument38 pagesOrganic AcidMuhammad Adeel JavedNo ratings yet
- ABT 227 - Course Outline Introduction To Molecular Biology (3335)Document1 pageABT 227 - Course Outline Introduction To Molecular Biology (3335)justevansiNo ratings yet
- MSC MPlil ZoologyDocument18 pagesMSC MPlil ZoologyYougesh KumarNo ratings yet
- Culture of Bone Marrow CellsDocument40 pagesCulture of Bone Marrow CellsMunesh SherawatNo ratings yet
- Bio Edit Software ReviewDocument3 pagesBio Edit Software ReviewNathalia Clavijo50% (2)
- BS Zoology: 5th Semester Placentation in MammalsDocument30 pagesBS Zoology: 5th Semester Placentation in MammalsTask BirdNo ratings yet
- 11th Biology Question BankDocument14 pages11th Biology Question BankJay senthilNo ratings yet
- Gregor MendelDocument3 pagesGregor MendelhdNo ratings yet
- Flowering Plants Sexual ReproductionDocument12 pagesFlowering Plants Sexual ReproductionNishtha JainNo ratings yet
- TSSM Unit Two Exam 2017Document19 pagesTSSM Unit Two Exam 2017nisulNo ratings yet
- Avatics 2017 03-02-15!34!57protein Synthesis Worksheet PracticeDocument2 pagesAvatics 2017 03-02-15!34!57protein Synthesis Worksheet PracticeMiguel BernalNo ratings yet
- 2 - Cell Culture LectureDocument45 pages2 - Cell Culture LectureSathvik PalakurtyNo ratings yet
- Protein Extraction From Lupin Seeds-A Mathematical ModelDocument11 pagesProtein Extraction From Lupin Seeds-A Mathematical Modeltbbdocument1s0% (1)
- Bacterial Ghost As Delivery SystemDocument49 pagesBacterial Ghost As Delivery SystemSalma El Faham100% (1)
- Cell Division, Mitosis, and CancerDocument25 pagesCell Division, Mitosis, and CancerChristopher PhillipsNo ratings yet
- Bacterial Antigenic StructureDocument36 pagesBacterial Antigenic Structuredearsaran0% (1)
- Fungi Morphology and Classification ExplainedDocument37 pagesFungi Morphology and Classification Explainedjihan faradhilaNo ratings yet
- Intermediate FilamentsDocument4 pagesIntermediate FilamentsSai SridharNo ratings yet
- Micrology and Parasitology ReviewerDocument44 pagesMicrology and Parasitology Reviewermarjourie cadampogNo ratings yet
- Translation and Proteins: The Products of TranscriptionDocument10 pagesTranslation and Proteins: The Products of TranscriptionRenz L. SalumbreNo ratings yet
- Biochemistry IIDocument2 pagesBiochemistry IISanthosh Kalash100% (1)
- Experiment 9 Bioinformatics Tools For Cell and Molecular BiologyDocument11 pagesExperiment 9 Bioinformatics Tools For Cell and Molecular BiologyJedd VirgoNo ratings yet
- LSM3233 Lecture 1Document43 pagesLSM3233 Lecture 1Prab NathanNo ratings yet
- Endocytosis ExocytosisDocument5 pagesEndocytosis ExocytosisMiska Fairuz100% (1)
- Wiley - Gene Cloning and DNA Analysis An Introduction, 7th Edition - 978-1-119-07256-0Document2 pagesWiley - Gene Cloning and DNA Analysis An Introduction, 7th Edition - 978-1-119-07256-0alfiNo ratings yet
- Lab 09 HandoutDocument8 pagesLab 09 HandoutHyenaNo ratings yet
- Cytology McqsDocument8 pagesCytology McqsZiyad AbdallahNo ratings yet
- SerialDilutions PDFDocument5 pagesSerialDilutions PDFAmelia_KharismayantiNo ratings yet
- GCSE Biology - Mitosis and Meiosis Spies and TrainsDocument8 pagesGCSE Biology - Mitosis and Meiosis Spies and TrainsTamicka BonnickNo ratings yet
- Physiology of BacteriaDocument150 pagesPhysiology of BacteriaМохіт Кумар ЯмпатіNo ratings yet
- Mitosis and Meiosis Discussion QuestionsDocument5 pagesMitosis and Meiosis Discussion QuestionsRichard Rna NaabNo ratings yet
- Tumores Solitarios Pulmonares InusualesDocument8 pagesTumores Solitarios Pulmonares InusualesSasha de la CruzNo ratings yet
- The Molecular Basis of Sperm-Oocyte Membrane Interactions During Mammalian FertilizationDocument0 pagesThe Molecular Basis of Sperm-Oocyte Membrane Interactions During Mammalian FertilizationSasha de la CruzNo ratings yet
- Understanding The Molecular Basis of Sperm Capacitation Through Kinase DesignDocument2 pagesUnderstanding The Molecular Basis of Sperm Capacitation Through Kinase DesignSasha de la CruzNo ratings yet
- Mammalian Fertilization - Molecular Aspects of Gamete Adhesion, Exocytosis and FusionDocument9 pagesMammalian Fertilization - Molecular Aspects of Gamete Adhesion, Exocytosis and FusionSasha de la CruzNo ratings yet
- Signalling Pathways Involved in Sperm CapacitationDocument0 pagesSignalling Pathways Involved in Sperm CapacitationSasha de la CruzNo ratings yet
- Señales Moleculares de Hipertrofia CardíacaDocument8 pagesSeñales Moleculares de Hipertrofia CardíacaSasha de la CruzNo ratings yet
- Clin Infect Dis.-2013-Wallis-106-13 PDFDocument8 pagesClin Infect Dis.-2013-Wallis-106-13 PDFSasha de la CruzNo ratings yet
- Clin Infect Dis.-2012 - 1228-35 PDFDocument8 pagesClin Infect Dis.-2012 - 1228-35 PDFSasha de la CruzNo ratings yet
- Infecciones Helmínticas PulmonaresDocument11 pagesInfecciones Helmínticas PulmonaresSasha de la CruzNo ratings yet
- Aspergilosis Pulmonares No InvasivasDocument21 pagesAspergilosis Pulmonares No InvasivasSasha de la CruzNo ratings yet
- Regulacion de La Produccion Del SurfactanteDocument11 pagesRegulacion de La Produccion Del SurfactanteSasha de la CruzNo ratings yet
- Patogenia de La Toxoplasmosis OcularDocument4 pagesPatogenia de La Toxoplasmosis OcularSasha de la CruzNo ratings yet
- Patogenia de La ToxoplasmosisDocument10 pagesPatogenia de La ToxoplasmosisSasha de la CruzNo ratings yet
- Patogenia de La GiardiasisDocument5 pagesPatogenia de La GiardiasisSasha de la CruzNo ratings yet
- Patogenia de Las Infecciones Por AcanthamoebaDocument9 pagesPatogenia de Las Infecciones Por AcanthamoebaSasha de la CruzNo ratings yet
- Patogenia de La AmebiasisDocument5 pagesPatogenia de La AmebiasisSasha de la CruzNo ratings yet
- Aspergilosis - Diagnóstico y TratamientoDocument8 pagesAspergilosis - Diagnóstico y TratamientoSasha de la CruzNo ratings yet
- Lipofuscinosis Ceroidea Neuronal - IIDocument6 pagesLipofuscinosis Ceroidea Neuronal - IISasha de la CruzNo ratings yet
- Anisakiosis - IIDocument5 pagesAnisakiosis - IISasha de la CruzNo ratings yet
- Clinical Review The Management of Hypertensive CrisesDocument11 pagesClinical Review The Management of Hypertensive CrisesAdam KurniaNo ratings yet
- Hypertensive CrisisDocument13 pagesHypertensive Crisis.Katherine CalderonNo ratings yet
- Crisis Hipertensivas - IVDocument5 pagesCrisis Hipertensivas - IVSasha de la CruzNo ratings yet
- Obesidad y Asma - IDocument7 pagesObesidad y Asma - ISasha de la CruzNo ratings yet
- Eh ChestDocument16 pagesEh ChestEdizellNo ratings yet
- Espectro Autista - IIDocument12 pagesEspectro Autista - IISasha de la CruzNo ratings yet
- Lipofuscinosis Ceroidea Neuronal - IDocument4 pagesLipofuscinosis Ceroidea Neuronal - ISasha de la CruzNo ratings yet
- Does The Ovum Select Her Gender?: - DR Muhammad El Hennawy - Ob/gyn SpecialistDocument18 pagesDoes The Ovum Select Her Gender?: - DR Muhammad El Hennawy - Ob/gyn Specialistkelapamuda21No ratings yet
- G5 Science - Q1-W5 - Male Reproductive SystemDocument20 pagesG5 Science - Q1-W5 - Male Reproductive SystemMarimel GuillenaNo ratings yet
- Chapter 7 FertilizationDocument44 pagesChapter 7 Fertilizationyacla1No ratings yet
- Exemplo - Reproduction, Fertility and DevelopmentDocument8 pagesExemplo - Reproduction, Fertility and DevelopmentMaiko DantasNo ratings yet
- Human Embryology StagesDocument9 pagesHuman Embryology StagesAdvin BurkeNo ratings yet
- Antifertility Effect of Costus Lucanuscianus Stem Extract in Male Albino RatsDocument5 pagesAntifertility Effect of Costus Lucanuscianus Stem Extract in Male Albino Ratsvictoria obinnaNo ratings yet
- Gnetum GnemonDocument15 pagesGnetum GnemonmastikaNo ratings yet
- Aves - Polispermia Supervivencia EmbrionariaDocument6 pagesAves - Polispermia Supervivencia EmbrionariaMarco AsprónNo ratings yet
- Biology Form 5 Chapter 4 QuestionsDocument12 pagesBiology Form 5 Chapter 4 QuestionsBoyd Junior67% (3)
- Oxo Act01 b303 As01 XxaannDocument3 pagesOxo Act01 b303 As01 XxaannMaisieNo ratings yet
- Sperm MorphologyDocument4 pagesSperm MorphologyDr.Jignesh chaudharyNo ratings yet
- Semen AnalysisDocument1 pageSemen AnalysisReyn BalbuenaNo ratings yet
- f3 - Notes 4.1 4 - ReproductionDocument12 pagesf3 - Notes 4.1 4 - ReproductionhermanttfNo ratings yet
- Genetics Concept and DefinitionDocument90 pagesGenetics Concept and DefinitionsomiNo ratings yet
- W6-8-Understanding The SelfDocument19 pagesW6-8-Understanding The Selfiser kyle LabanderoNo ratings yet
- June 2012 MS - Paper 1B Edexcel Biology IGCSEDocument18 pagesJune 2012 MS - Paper 1B Edexcel Biology IGCSESadhanshyhan ShyhanNo ratings yet
- Igcse BioDocument17 pagesIgcse BioimeNo ratings yet
- Sexual Reproduction Grade 10Document23 pagesSexual Reproduction Grade 10Carol EscañoNo ratings yet
- Module 1 Class XII - Biology EFDocument144 pagesModule 1 Class XII - Biology EFpadmaNo ratings yet
- Blighted Ovum Anembryonic Pregnancy: Mary M. Gallenberg, M.DDocument4 pagesBlighted Ovum Anembryonic Pregnancy: Mary M. Gallenberg, M.Dadminbunda sehatNo ratings yet
- Gametogenesis: The Production of Eggs and SpermDocument15 pagesGametogenesis: The Production of Eggs and SpermvimamufliantiNo ratings yet
- How Do Organisms Reproduce Important Questions PDFDocument12 pagesHow Do Organisms Reproduce Important Questions PDFSomik JainNo ratings yet
- About Lobsters 1962Document196 pagesAbout Lobsters 1962ana_petrescuNo ratings yet
- Forensic Science Multiple Choice QuestionsDocument3 pagesForensic Science Multiple Choice QuestionsAra Joyce IINo ratings yet
- Semen analysis report title under 40 charsDocument10 pagesSemen analysis report title under 40 charsraghava kulkarniNo ratings yet