You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Mission S and W Dims and Curve Chart PDFDocument28 pagesMission S and W Dims and Curve Chart PDFcaballero_tigreNo ratings yet
- TDS-Dow SPECFIL FT630 & SPECFIL FE100-EN - 20181226Document2 pagesTDS-Dow SPECFIL FT630 & SPECFIL FE100-EN - 20181226Mallampati RamakrishnaNo ratings yet
- Gas Natural TransmisionDocument43 pagesGas Natural Transmisionangel3reyesNo ratings yet
- Mixing TimeDocument7 pagesMixing TimeakoNo ratings yet
- Biochemical Engineering and Biotechnology Handbook ATKINSONDocument568 pagesBiochemical Engineering and Biotechnology Handbook ATKINSONMonique Lopes da Silva100% (4)
- Innovation of LNGCDocument63 pagesInnovation of LNGCpadangiringNo ratings yet
- Ácidos Polipróticos (Espécies Intermediárias)Document7 pagesÁcidos Polipróticos (Espécies Intermediárias)Monique Lopes da SilvaNo ratings yet
- Thiuram MixDocument4 pagesThiuram MixMonique Lopes da SilvaNo ratings yet
- 13 Secrets For Speaking Fluent Japanese PDFDocument96 pages13 Secrets For Speaking Fluent Japanese PDFMonique Lopes da SilvaNo ratings yet
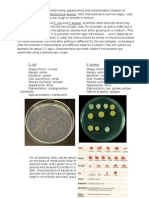
- Bacteria Colony Appearance MorphologyDocument2 pagesBacteria Colony Appearance MorphologyMonique Lopes da SilvaNo ratings yet
- A Critical Review of The Methods For Cleavage of Fusion ProteinsDocument11 pagesA Critical Review of The Methods For Cleavage of Fusion ProteinsMonique Lopes da SilvaNo ratings yet
- Medical Microbiology: Corynebacterium SpeciesDocument6 pagesMedical Microbiology: Corynebacterium SpeciesMonique Lopes da SilvaNo ratings yet
- Sensors Evaluation 2019 Oct 19 (19) - DOC#NDL:NDA:DGA:ON:072002 PDFDocument1 pageSensors Evaluation 2019 Oct 19 (19) - DOC#NDL:NDA:DGA:ON:072002 PDFDivyansh KohliNo ratings yet
- Dr.K. Prabha PDFDocument4 pagesDr.K. Prabha PDFVrsarma DhulipalaNo ratings yet
- Preparation and Properties of Compounds - 01-TheoryDocument45 pagesPreparation and Properties of Compounds - 01-TheoryRaju SinghNo ratings yet
- Chapter 9 The Endocrine SystemDocument1 pageChapter 9 The Endocrine SystemacNo ratings yet
- Hacon TMDocument68 pagesHacon TMTiago CoutoNo ratings yet
- ECSS Q ST 70 15C (1may2021)Document124 pagesECSS Q ST 70 15C (1may2021)Navamani Prakash100% (1)
- Florgard Epu SL - TDSDocument3 pagesFlorgard Epu SL - TDSGabriel GabeNo ratings yet
- Metabolism Exam 2 - GIFT - Spring 2016Document9 pagesMetabolism Exam 2 - GIFT - Spring 2016shafa_nathani100% (2)
- David Scanu, Natalia P. Yevlampieva and Robert Deschenaux - Polar and Electrooptical Properties of (60) Fullerene-Containing Poly (Benzyl Ether) Dendrimers in SolutionDocument7 pagesDavid Scanu, Natalia P. Yevlampieva and Robert Deschenaux - Polar and Electrooptical Properties of (60) Fullerene-Containing Poly (Benzyl Ether) Dendrimers in SolutionGlade680No ratings yet
- Nuclear Technology and EnergyDocument4 pagesNuclear Technology and EnergyJessica Anna LuchkaNo ratings yet
- Stabilisation of Soft Clay Using Ground Granulated Blastfurnace Slag and LimeDocument7 pagesStabilisation of Soft Clay Using Ground Granulated Blastfurnace Slag and LimeVivek SvkNo ratings yet
- 5070 s23 QP 42 MergedDocument100 pages5070 s23 QP 42 MergedMuhammad QasimNo ratings yet
- Rhododendron Anthopogon 3Document11 pagesRhododendron Anthopogon 3subashrai7991No ratings yet
- BIO Advancing The Biobased Economy 2016Document84 pagesBIO Advancing The Biobased Economy 2016unicornmfkNo ratings yet
- 202 Introduction To Materials ScienceDocument435 pages202 Introduction To Materials ScienceRK SinghNo ratings yet
- Naufal - Flare Technologies - PPTX (Tanpa Video)Document12 pagesNaufal - Flare Technologies - PPTX (Tanpa Video)Naufal AlhadyNo ratings yet
- Oxidation-Reduction of Borneol PDFDocument8 pagesOxidation-Reduction of Borneol PDFRalston King Stulla ChambersNo ratings yet
- Tutorial ERP StudentsDocument2 pagesTutorial ERP StudentsHariz MiorNo ratings yet
- Growth and Characterization of L-Alanine Potassium Nitrate Single Crystals For Nonlinear Optical ApplicationsDocument5 pagesGrowth and Characterization of L-Alanine Potassium Nitrate Single Crystals For Nonlinear Optical ApplicationsPalaniswamy SankariahNo ratings yet
- A Case Study On Solar Vapour Absorption Refrigeration SystemDocument7 pagesA Case Study On Solar Vapour Absorption Refrigeration SystemShivam MaheraNo ratings yet
- Cod and BodDocument57 pagesCod and Boddinesh kumarNo ratings yet
- Energy BalanceDocument28 pagesEnergy BalanceEian HawNo ratings yet
- Presentation: Darwisa G. Hawari Dip-Ems 1ADocument35 pagesPresentation: Darwisa G. Hawari Dip-Ems 1AZeal Alexis MontellorNo ratings yet
- Experiment NoDocument5 pagesExperiment NoAbdullah UsmaniNo ratings yet
- Inviscid ProblemsDocument6 pagesInviscid Problemssoe0303No ratings yet