You might also like
- Seminar: Sakib Burza, Simon L Croft, Marleen BoelaertDocument20 pagesSeminar: Sakib Burza, Simon L Croft, Marleen BoelaertMarco Antonio KoffNo ratings yet
- Experimental Parasitology: A. Zabala-Peñafiel, D. Todd, H. Daneshvar, R. Burchmore TDocument7 pagesExperimental Parasitology: A. Zabala-Peñafiel, D. Todd, H. Daneshvar, R. Burchmore TLeidy Tatiana Merchán RoaNo ratings yet
- Jurnal Kel. 2 FilariasisDocument9 pagesJurnal Kel. 2 FilariasisEka Saptaria NusantiNo ratings yet
- Leishmaniasis Fact Sheet N 375Document6 pagesLeishmaniasis Fact Sheet N 375mclimacoNo ratings yet
- Solomon ZoologyDocument29 pagesSolomon ZoologyYARO TERKIMBINo ratings yet
- Immunity To Visceral Leishmaniasis Using Genetically Defined Live-Attenuated ParasitesDocument13 pagesImmunity To Visceral Leishmaniasis Using Genetically Defined Live-Attenuated ParasitesmclimacoNo ratings yet
- Othman 2014Document44 pagesOthman 2014Jessica QuevedoNo ratings yet
- LeishmaniasisDocument7 pagesLeishmaniasisLuis Carlos Quinto CuzcanoNo ratings yet
- Abp 2 - Leishmaniasis: Key FactsDocument4 pagesAbp 2 - Leishmaniasis: Key FactsNatalia ChauNo ratings yet
- Leishmania SisDocument9 pagesLeishmania SisAli khanNo ratings yet
- Tropical Diseases An Unsolved ChallengeDocument9 pagesTropical Diseases An Unsolved ChallengeYet Barreda BasbasNo ratings yet
- 2511 5848 1 PB PDFDocument12 pages2511 5848 1 PB PDFDewi SiregarNo ratings yet
- 2511 5848 1 PB PDFDocument12 pages2511 5848 1 PB PDFeva yustianaNo ratings yet
- Wahidspaper 1Document61 pagesWahidspaper 1راوند اعبيدNo ratings yet
- Mechanisms of Immunopathogenesis 2021Document16 pagesMechanisms of Immunopathogenesis 2021MARIE SELENE RAMIREZ REVOLLARNo ratings yet
- Leishmaniasis, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsFrom EverandLeishmaniasis, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsNo ratings yet
- A Review of Leishmaniasis: Current Knowledge and Future DirectionsDocument12 pagesA Review of Leishmaniasis: Current Knowledge and Future DirectionsJaison Enrique Torres PachecoNo ratings yet
- LeishmaniasisDocument17 pagesLeishmaniasisAkram RashidNo ratings yet
- Leishmaniasis: Key FactsDocument6 pagesLeishmaniasis: Key FactsNurul Hikmah PratiwiNo ratings yet
- Review of S.Aureus in Cat WoundsDocument20 pagesReview of S.Aureus in Cat WoundsM.Aleem khanNo ratings yet
- Genetic Approaches For Malaria Control: Marcelo Jacobs-LorenaDocument14 pagesGenetic Approaches For Malaria Control: Marcelo Jacobs-Lorenayenniffer VicenteNo ratings yet
- Proquest Jurnal 7Document19 pagesProquest Jurnal 7Winda PNo ratings yet
- Leishmaniasis: A Review (Version 1 Peer Review: 2 Approved)Document15 pagesLeishmaniasis: A Review (Version 1 Peer Review: 2 Approved)odhiambo samwel100% (1)
- ©201 1 Landes Bi Osci Ence. Do Not Di ST R I But E.: LeishmaniasisDocument11 pages©201 1 Landes Bi Osci Ence. Do Not Di ST R I But E.: LeishmaniasisNovericko Ginger BudionoNo ratings yet
- 2018 Leishmania Meta Analysis in EthiopiaDocument23 pages2018 Leishmania Meta Analysis in EthiopiafedhawakNo ratings yet
- The Antimicrobial Resistance Crisis: Causes, Consequences, and ManagementDocument8 pagesThe Antimicrobial Resistance Crisis: Causes, Consequences, and ManagementPichayaNo ratings yet
- HTTPS:WWW Ncbi NLM Nih gov:pmc:articles:PMC9867007:pdf:viruses-15-00225Document17 pagesHTTPS:WWW Ncbi NLM Nih gov:pmc:articles:PMC9867007:pdf:viruses-15-00225Don SarNo ratings yet
- Nyas - Streit and Lafontant 7081425036Document11 pagesNyas - Streit and Lafontant 7081425036Thomas StreitNo ratings yet
- Genome: Campylobacter (Meaning 'Twisted Bacteria') Is A Genus of Bacteria That Are Gram-Negative, Spiral, andDocument7 pagesGenome: Campylobacter (Meaning 'Twisted Bacteria') Is A Genus of Bacteria That Are Gram-Negative, Spiral, andArsh McBarsh MalhotraNo ratings yet
- Tansboundary Animal Diseases A ReviewDocument58 pagesTansboundary Animal Diseases A ReviewSohaib AslamNo ratings yet
- Veterinary Clinical Pathology Clerkship ProgramDocument46 pagesVeterinary Clinical Pathology Clerkship ProgramDrVijayata ChoudharyNo ratings yet
- MALARIA Descriptive Epidemiology of MalariaDocument16 pagesMALARIA Descriptive Epidemiology of MalariaAms BeeNo ratings yet
- Malaria 2018Document11 pagesMalaria 2018gerges8723No ratings yet
- Literature Review SalmonellaDocument7 pagesLiterature Review Salmonellaafdtbwkhb100% (1)
- Lassa Fever PaperDocument18 pagesLassa Fever PaperCherica OnateNo ratings yet
- Review of Leishmaniasis in The Middle East and North AfricaDocument9 pagesReview of Leishmaniasis in The Middle East and North Africameryem.assidiNo ratings yet
- Leishmania Synthesis PaperDocument7 pagesLeishmania Synthesis Paperkristiaa_1No ratings yet
- Knowledge Attutide and Practice of Ameobiasis Among Mothers in Washinton Idp Garowe Puntland SomaliaDocument6 pagesKnowledge Attutide and Practice of Ameobiasis Among Mothers in Washinton Idp Garowe Puntland SomaliaAbdirahman YazinNo ratings yet
- LEPTOSPIROSIS ReviewDocument10 pagesLEPTOSPIROSIS ReviewwaktoleNo ratings yet
- 01 AEXTJ 389 23 RevisedDocument6 pages01 AEXTJ 389 23 RevisedBRNSS Publication Hub InfoNo ratings yet
- Ijarbs 2Document23 pagesIjarbs 2Sajjad LaghariNo ratings yet
- ADOLE M.SC FinalDocument61 pagesADOLE M.SC FinalTerna HonNo ratings yet
- Pathogens 12 00776Document2 pagesPathogens 12 00776Hashan AbeyrathneNo ratings yet
- LSDV Synopsis 1Document13 pagesLSDV Synopsis 1Bilal RaheemNo ratings yet
- Article1380870598 - Pérez Et AlDocument8 pagesArticle1380870598 - Pérez Et AlnepretipNo ratings yet
- Review On Epidemiology and Pathology of Yellow Fever VirusDocument24 pagesReview On Epidemiology and Pathology of Yellow Fever Virushenok bushura100% (1)
- Malaria Eradication A War How To Win PDFDocument6 pagesMalaria Eradication A War How To Win PDFashishNo ratings yet
- Kelompok 1 Leishmaniasis-A-Review-On-Parasite-Vector-And-Reservoir-HostDocument6 pagesKelompok 1 Leishmaniasis-A-Review-On-Parasite-Vector-And-Reservoir-HostPanji SetyawanNo ratings yet
- SalmonellaDocument14 pagesSalmonellaTochukwu IbekweNo ratings yet
- Is HIV The Biggest Mistake of XXcenturyDocument14 pagesIs HIV The Biggest Mistake of XXcenturyMartini100% (1)
- Economic Effect Parasite Assignment 3Document4 pagesEconomic Effect Parasite Assignment 3MUHAMMADNo ratings yet
- Aftab,+28 the+Economic+Impact+of+Lumpy+Skin+DiseaseDocument7 pagesAftab,+28 the+Economic+Impact+of+Lumpy+Skin+DiseaseHanif Ur RahmanNo ratings yet
- A Review On Bovine Brucellosis: Epidemiology, Diagnosis and Control OptionsDocument14 pagesA Review On Bovine Brucellosis: Epidemiology, Diagnosis and Control OptionsSajjad LaghariNo ratings yet
- Evolutionary Perspectives On Wildlife Disease: Concepts and ApplicationsDocument8 pagesEvolutionary Perspectives On Wildlife Disease: Concepts and ApplicationsKaritoNo ratings yet
- Control of Neglected Tropical Diseases: Review ArticleDocument10 pagesControl of Neglected Tropical Diseases: Review ArticleDewi Puspita ApsariNo ratings yet
- Running Head: MALARIA 1Document7 pagesRunning Head: MALARIA 1klm klmNo ratings yet
- Rehema Research ProposalDocument8 pagesRehema Research Proposalwillisonjohn155No ratings yet
- A Review Therapeutics AmebiasisDocument9 pagesA Review Therapeutics AmebiasisSandrina NajwaNo ratings yet
- Tuberculosis in Pregnancy Students' LectureDocument13 pagesTuberculosis in Pregnancy Students' LectureAsri Indriyani PutriNo ratings yet
- 2020-COR-TXT Planetary Emergency 2-0-Aug2020Document9 pages2020-COR-TXT Planetary Emergency 2-0-Aug2020Mika DeverinNo ratings yet
- Traumatic Tympanic PerforationDocument14 pagesTraumatic Tympanic PerforationNatalia CvNo ratings yet
- ASHHC Patient Action Plan FPR COPD Signs and SymptomsDocument2 pagesASHHC Patient Action Plan FPR COPD Signs and SymptomsOscar CocNo ratings yet
- Summative TestDocument4 pagesSummative TestGlenda GlendaNo ratings yet
- Gram Negative SpirochetesDocument50 pagesGram Negative SpirochetesYeshiwas FelekeNo ratings yet
- WA 2009 Certificate of ExemptionDocument1 pageWA 2009 Certificate of ExemptionDonnaNo ratings yet
- HydrotherapyDocument26 pagesHydrotherapyMohd Ismarezza100% (2)
- Name of Drug SoludexideDocument2 pagesName of Drug SoludexideSian AsadaNo ratings yet
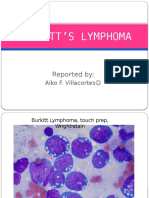
- Burkitt S LymphomaDocument12 pagesBurkitt S LymphomaAiko VillacortesNo ratings yet
- First Internal Assessment Political Science III International RelationsDocument2 pagesFirst Internal Assessment Political Science III International RelationsUjjwal AnandNo ratings yet
- Part A-8-Proving Child Survival RatesDocument7 pagesPart A-8-Proving Child Survival RatesdarylNo ratings yet
- Chapter 1 Science Form 1Document19 pagesChapter 1 Science Form 1Che SalNo ratings yet
- A Short History of The Stamford & Rutland HospitalDocument47 pagesA Short History of The Stamford & Rutland HospitalJohn McKechnie100% (1)
- Michaels, Paula - Medical Propaganda and Cultural RevolutionDocument21 pagesMichaels, Paula - Medical Propaganda and Cultural RevolutionAinura ToksanbayevaNo ratings yet
- CHN Post Test Students Copy NewDocument3 pagesCHN Post Test Students Copy NewMeeKo Videña100% (1)
- TetrapareseDocument38 pagesTetraparesemariamunsriNo ratings yet
- Crisis of LeadershipDocument1 pageCrisis of LeadershipGC MachineNo ratings yet
- GF 689 Poverty and HealthDocument4 pagesGF 689 Poverty and HealthTowardinho14No ratings yet
- Avenue of PenetrationDocument3 pagesAvenue of Penetrationshyamsunder68100% (1)
- Mid TermDocument17 pagesMid TermTamoghna NaskarNo ratings yet
- Health Education DHFDocument19 pagesHealth Education DHFIshaBrijeshSharma100% (5)
- Bacillus Anthracis: Basic Laboratory Protocols For The Presumptive Identification ofDocument16 pagesBacillus Anthracis: Basic Laboratory Protocols For The Presumptive Identification ofMyro DarlandanNo ratings yet
- HaemoparasitesDocument82 pagesHaemoparasitesAshok WiselinNo ratings yet
- IGCSE English Language Revision GuideDocument3 pagesIGCSE English Language Revision GuideAjay LakshmananNo ratings yet
- Who CDS Epr Lyo 2006 2 PDFDocument90 pagesWho CDS Epr Lyo 2006 2 PDFChyntiaDRNo ratings yet
- BACTERIOPHAGEDocument11 pagesBACTERIOPHAGEDivyanshu YadavNo ratings yet
- Acute Gingival InfectionsDocument35 pagesAcute Gingival InfectionsAhmed Tawfig GamalNo ratings yet
- Grade 10 - Q1 - W5 L1 - Compare and ContrastDocument7 pagesGrade 10 - Q1 - W5 L1 - Compare and ContrastAndre Marell Cacatian100% (1)
- Nursing Care Plan For Pneumonia NCPDocument3 pagesNursing Care Plan For Pneumonia NCPFARAH MAE MEDINANo ratings yet