You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Abdrehim y RepoooDocument8 pagesAbdrehim y RepoooIsabel ImanNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- 59 2 3Document4 pages59 2 3Isabel ImanNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Abderrahim2015 1 PDFDocument8 pagesAbderrahim2015 1 PDFConsuelo VegaNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Betalaina FitoquimicaDocument29 pagesBetalaina FitoquimicaIsabel ImanNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Fibra DietariaDocument6 pagesFibra DietariaIsabel ImanNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- 1 s2.0 S0092867407013402 MainDocument11 pages1 s2.0 S0092867407013402 MainIsabel ImanNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Swain1959 Metodología para Fenolicos Totales PDFDocument6 pagesSwain1959 Metodología para Fenolicos Totales PDFIsabel ImanNo ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Determination of Total Antioxidant Capacity in Kañihua Paper Boliviano PDFDocument5 pagesDetermination of Total Antioxidant Capacity in Kañihua Paper Boliviano PDFIsabel ImanNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Repo Composicion Nutrcional de CañihuaDocument12 pagesRepo Composicion Nutrcional de CañihuaIsabel ImanNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Flavonoides Springer 2Document10 pagesFlavonoides Springer 2Isabel ImanNo ratings yet
- Repo y Valencia Cañihua Grano y Pan PDFDocument8 pagesRepo y Valencia Cañihua Grano y Pan PDFIsabel ImanNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Campos 2006Document8 pagesCampos 2006Isabel ImanNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- BETALAINAS Mattila2005Document6 pagesBETALAINAS Mattila2005Isabel ImanNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Paper KañihuaDocument10 pagesPaper KañihuaIsabel ImanNo ratings yet
- Abts AssayDocument7 pagesAbts AssayKadashaNo ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Betalainas en QuinuaDocument8 pagesBetalainas en QuinuaIsabel ImanNo ratings yet
- Flavonoides Springer 2Document10 pagesFlavonoides Springer 2Isabel ImanNo ratings yet
- CULTURALZOODocument5 pagesCULTURALZOOIsabel ImanNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Cultural ZooDocument5 pagesCultural ZooIsabel ImanNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Elevated Intz Tank 279Document23 pagesElevated Intz Tank 279Navasivayam Sankar100% (2)
- BEP Pages 5 16Document12 pagesBEP Pages 5 16Amy LauNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Dry ContactsDocument2 pagesDry ContactsDiwakar MishraNo ratings yet
- Flextronics FINALDocument25 pagesFlextronics FINALAnkit GauravNo ratings yet
- Designs of Canals and CM&CD WorksDocument61 pagesDesigns of Canals and CM&CD WorksVenkataLakshmiKorrapatiNo ratings yet
- Cutting Temperature - Causes - Effects - Estimation (Assessment) & ControlDocument14 pagesCutting Temperature - Causes - Effects - Estimation (Assessment) & ControlvelladuraiNo ratings yet
- Cryogenics Handbook PDFDocument227 pagesCryogenics Handbook PDFmangyanNo ratings yet
- Didsn1811a (Skyair R-410a)Document12 pagesDidsn1811a (Skyair R-410a)Ari SetyawanNo ratings yet
- Blackboard 9.1Document49 pagesBlackboard 9.1ds532No ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Lab 3 SST 4303Document5 pagesLab 3 SST 4303Muazrul MangsorNo ratings yet
- Business Plan On Plastic Recycling & ManufacturingDocument19 pagesBusiness Plan On Plastic Recycling & ManufacturingSah Sanjiv KumarNo ratings yet
- EBS Data MaskingDocument31 pagesEBS Data MaskingNawab ShariefNo ratings yet
- BoyapatyDocument5 pagesBoyapatylalalallalalaaluuuNo ratings yet
- Continuous Thread Stud Double End Stud ADocument11 pagesContinuous Thread Stud Double End Stud AMarius Mihai Buzduga0% (1)
- Cryptographic Systems Tend To Involve Both Algorithm and ADocument6 pagesCryptographic Systems Tend To Involve Both Algorithm and APawan Kumar ThakurNo ratings yet
- Noise and VibrationDocument14 pagesNoise and VibrationABHISHEK V GOWDANo ratings yet
- Catalogo CMCDocument145 pagesCatalogo CMCalyboscanNo ratings yet
- Annex12 Technical Cards For Accessible ConstructionDocument44 pagesAnnex12 Technical Cards For Accessible Constructionarchitectfemil6663No ratings yet
- AIP Puducherry Aerodrome, IndiaDocument13 pagesAIP Puducherry Aerodrome, Indiass khadriNo ratings yet
- Robotics Engineering Minor FlowchartDocument1 pageRobotics Engineering Minor FlowchartkskkingNo ratings yet
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Resistor DatasheetDocument10 pagesResistor DatasheetEndradno KurniaNo ratings yet
- Standards of MeasurementDocument12 pagesStandards of MeasurementShubham KheraNo ratings yet
- A Control System For DC Arc Furnaces For Power Quality ImprovementsDocument8 pagesA Control System For DC Arc Furnaces For Power Quality ImprovementsgtgreatNo ratings yet
- 010 Zircoat MDocument3 pages010 Zircoat MvsdeepakchandraNo ratings yet
- Analysis Procedure K2co3 PDFDocument3 pagesAnalysis Procedure K2co3 PDFPiyush PatelNo ratings yet
- NextCloud OnlyOffice PDFDocument89 pagesNextCloud OnlyOffice PDFOmar Gallegos Vazquez100% (1)
- BS en 50483 6 2009Document27 pagesBS en 50483 6 2009Shara LogisticNo ratings yet
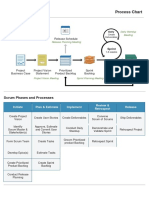
- Procesos SCRUMDocument2 pagesProcesos SCRUMRoseNo ratings yet
- Castrol - Iceomatic SW 68Document7 pagesCastrol - Iceomatic SW 68advantage025No ratings yet
- EnglishDocument6 pagesEnglishThineswary Siva naiduNo ratings yet
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Periodic Tales: A Cultural History of the Elements, from Arsenic to ZincFrom EverandPeriodic Tales: A Cultural History of the Elements, from Arsenic to ZincRating: 3.5 out of 5 stars3.5/5 (137)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)