You might also like
- Food Biotechnology: Fabia YousafDocument6 pagesFood Biotechnology: Fabia Yousaffabia yousafNo ratings yet
- Lipases in Dairy Industry ReviewDocument10 pagesLipases in Dairy Industry ReviewChinh Đinh ViệtNo ratings yet
- Enzyme ProductionDocument9 pagesEnzyme ProductionSweettha GunasekaranNo ratings yet
- Food Fermentation: Enzyme ProductionDocument9 pagesFood Fermentation: Enzyme ProductionSweettha GunasekaranNo ratings yet
- Food Fermentation - 23 Desember 2020 - FitDocument11 pagesFood Fermentation - 23 Desember 2020 - Fitlittle liarNo ratings yet
- 125897-Article Text-342726-1-10-20151116Document6 pages125897-Article Text-342726-1-10-20151116tonima rahmanNo ratings yet
- International College Food Production - DairyDocument11 pagesInternational College Food Production - DairyArina HazirahNo ratings yet
- Enzyme Production-Insdustrial MicrobiologyDocument7 pagesEnzyme Production-Insdustrial MicrobiologyPKSachanNo ratings yet
- Enzymes & Food Technology - ppt1Document32 pagesEnzymes & Food Technology - ppt1Abdullah AshrafNo ratings yet
- Microorganisms Role in AgricultureDocument41 pagesMicroorganisms Role in AgricultureMichael ThungNo ratings yet
- Role of biotechnology in food processing and preservationDocument51 pagesRole of biotechnology in food processing and preservationrani padamavathiNo ratings yet
- Fermented Foods IntroductionDocument7 pagesFermented Foods IntroductionAparajita GNo ratings yet
- Yeast Transgenic PlantsDocument5 pagesYeast Transgenic PlantsTooba Iqbal67% (6)
- Biotechnology in Microbial ProcessesDocument13 pagesBiotechnology in Microbial Processesolamicro50% (2)
- Bioproduccion de CompuestosDocument29 pagesBioproduccion de Compuestos3st3banruiz01No ratings yet
- How Microbes Help Create the Foods We LoveDocument27 pagesHow Microbes Help Create the Foods We LoveSwateja MangleNo ratings yet
- BiotechnologyDocument45 pagesBiotechnologycayananjefffrancisNo ratings yet
- MCB 407 Lectures Two and ThreeDocument8 pagesMCB 407 Lectures Two and ThreePopoola OlakunleNo ratings yet
- Midterm Enzyme Fermentation 1Document20 pagesMidterm Enzyme Fermentation 1Thuỳ TrangNo ratings yet
- Dairy Process Biotechnology DT-514Document34 pagesDairy Process Biotechnology DT-514aparnna vpNo ratings yet
- Microbiology: Fundamentals and Application: By: DR - Ir. Lilik Eka Radiati, MSDocument34 pagesMicrobiology: Fundamentals and Application: By: DR - Ir. Lilik Eka Radiati, MSSyira Zulkahfi Si KapiNo ratings yet
- Alternative Sources of Rennet for CheesemakingDocument2 pagesAlternative Sources of Rennet for CheesemakingZyla Monica JavierNo ratings yet
- Bio Tech DNADocument8 pagesBio Tech DNAWan NabilNo ratings yet
- Applications of Food BiotechnologyDocument4 pagesApplications of Food BiotechnologyAleksandra Sanja MartinovicNo ratings yet
- Enzymatic and Bacterial Conversions Duri PDFDocument10 pagesEnzymatic and Bacterial Conversions Duri PDFCiprian MocanNo ratings yet
- Ijair 3070 FinalDocument21 pagesIjair 3070 FinaldhokamokshaNo ratings yet
- Enzymes in Food Bioprocessing - Novel Food Enzymes, Applications, and Related TechniquesDocument6 pagesEnzymes in Food Bioprocessing - Novel Food Enzymes, Applications, and Related TechniquesRogelioSalazarNo ratings yet
- Future of Feed Enzymes: Orientation and Perspectives: R.R. MarquardtDocument7 pagesFuture of Feed Enzymes: Orientation and Perspectives: R.R. MarquardtdevivasantNo ratings yet
- Food IndustryDocument8 pagesFood IndustrySabnam HossainNo ratings yet
- Industrial ApplicationDocument25 pagesIndustrial Applicationphongbui100No ratings yet
- DK1209 CH02Document14 pagesDK1209 CH02Syed Ali Akbar BokhariNo ratings yet
- Microbial Enzymes for Dairy ProcessingDocument9 pagesMicrobial Enzymes for Dairy ProcessingrajadkvNo ratings yet
- Microbial Enzymes Improve Foods BeveragesDocument75 pagesMicrobial Enzymes Improve Foods BeveragesIna Ardan50% (2)
- Microbial Enzymes in Indian Food ProcessingDocument10 pagesMicrobial Enzymes in Indian Food ProcessingEshona ChakrabortyNo ratings yet
- Fermented sausage starter cultures effect on amino acidsDocument7 pagesFermented sausage starter cultures effect on amino acidssilviaNo ratings yet
- Unit 6 Biology Genetic Modification & MicroorganismDocument7 pagesUnit 6 Biology Genetic Modification & MicroorganismWendyNo ratings yet
- Diary Microbiology.9038206.PowerpointDocument18 pagesDiary Microbiology.9038206.PowerpointKIRUI JACKSONNo ratings yet
- Industrial Uses of Microbial EnzymesDocument23 pagesIndustrial Uses of Microbial EnzymesEhsan HumayunNo ratings yet
- Feed AdditiveDocument42 pagesFeed AdditiveNabeel ShahidNo ratings yet
- Biotech CropsDocument29 pagesBiotech CropsMalik UsamaNo ratings yet
- Capitulo 1 Bioprocess Engineering PrinciplesDocument9 pagesCapitulo 1 Bioprocess Engineering PrinciplesMariana Villegas BedoyaNo ratings yet
- BHM 3rd Food Sience and Nutrition Chapter 3Document9 pagesBHM 3rd Food Sience and Nutrition Chapter 3Suraj KhadkaNo ratings yet
- Bact EnzDocument42 pagesBact EnzKa QueenNo ratings yet
- Chapter-2 Enzyme Preparation and UseDocument33 pagesChapter-2 Enzyme Preparation and UsePreethy Thomas0% (2)
- Microbes in The Food Industry - Microorganisms - Biology: Fermented Dairy ProductsDocument7 pagesMicrobes in The Food Industry - Microorganisms - Biology: Fermented Dairy ProductsDeeksha MNo ratings yet
- Document: ArchivedDocument8 pagesDocument: ArchivedNaveenNo ratings yet
- Chapter 5 Microbial BiotechnologyDocument75 pagesChapter 5 Microbial BiotechnologySyamala Natarajan100% (1)
- Introduction of LipaseDocument38 pagesIntroduction of LipaseRamneet RanaNo ratings yet
- L. No. 1 Microbial EnzymesDocument3 pagesL. No. 1 Microbial EnzymesM Arfat YameenNo ratings yet
- Chapter 5 Microbial BiotechnologyDocument75 pagesChapter 5 Microbial BiotechnologymichaelNo ratings yet
- Food and Beverage BiotechnologyDocument10 pagesFood and Beverage BiotechnologyHayderNo ratings yet
- SBB 2203Document170 pagesSBB 2203Ram RamNo ratings yet
- Application of Biotechnology For The Biosynthesis of EnzymesDocument9 pagesApplication of Biotechnology For The Biosynthesis of EnzymesEstiarul PranNo ratings yet
- Economic Importance of FungiDocument13 pagesEconomic Importance of FungiskepticrelicNo ratings yet
- Food BiotechnologyDocument45 pagesFood Biotechnologyremosamy43No ratings yet
- Lecture 1Document20 pagesLecture 1warden tambiNo ratings yet
- Pertemuan - 13 Mikrobiologi Bahan MakananDocument25 pagesPertemuan - 13 Mikrobiologi Bahan MakananMahardika Adhi PratamaNo ratings yet
- Wa0013.Document50 pagesWa0013.Widya NingsiNo ratings yet
- Starter CulturesDocument11 pagesStarter CulturesMarcelina Mendoza SalazarNo ratings yet
- How To Mix Feed Rations With The Pearson Square: Grains, Protein, Calcium, Phosphorous, Balance, & MoreFrom EverandHow To Mix Feed Rations With The Pearson Square: Grains, Protein, Calcium, Phosphorous, Balance, & MoreNo ratings yet
- Extractie Cu CO2 Si Presare La ReceDocument11 pagesExtractie Cu CO2 Si Presare La ReceRazvan BabanNo ratings yet
- Extractie Cu CO2 Si Presare La ReceDocument11 pagesExtractie Cu CO2 Si Presare La ReceRazvan BabanNo ratings yet
- Application of Biosurfactants in Food Industry PDFDocument12 pagesApplication of Biosurfactants in Food Industry PDFRazvan BabanNo ratings yet
- Microbial Production of Natural FlavorsDocument9 pagesMicrobial Production of Natural FlavorsRazvan BabanNo ratings yet
- Common Authorisation ProcedureDocument30 pagesCommon Authorisation ProcedureRazvan BabanNo ratings yet
- CH 34Document27 pagesCH 34B6D4N0No ratings yet
- Immobilized β-Amylase StudyDocument7 pagesImmobilized β-Amylase StudyJhoana FalconNo ratings yet
- Microbial Production of Lactic Acid - The Latest DevelopmentDocument12 pagesMicrobial Production of Lactic Acid - The Latest DevelopmentMuhammad DliyauddinNo ratings yet
- Antimicrobial Activity of Alo e V e Ra (L.) Burm. F. Against Pathogenic MicroorganismsDocument8 pagesAntimicrobial Activity of Alo e V e Ra (L.) Burm. F. Against Pathogenic MicroorganismsBea NufuarNo ratings yet
- Englyst2000 StarchDocument19 pagesEnglyst2000 StarchNizel Shanin Noval NarsicoNo ratings yet
- Itaconic Acid Production by Filamentous Fungi in Starch-RichDocument7 pagesItaconic Acid Production by Filamentous Fungi in Starch-RichLaís CamposNo ratings yet
- Begin with Rice & Water: A Primer on Brewing MakgeolliDocument64 pagesBegin with Rice & Water: A Primer on Brewing MakgeolliNicholas Moy100% (1)
- Human A-Amylase and Starch Digestion: An Interesting MarriageDocument11 pagesHuman A-Amylase and Starch Digestion: An Interesting MarriagePratiksha RajpootNo ratings yet
- How enzymes work optimallyDocument111 pagesHow enzymes work optimallyCassey Hilario100% (4)
- Tdf100a Tdfc10 en Fibra DieteticaDocument4 pagesTdf100a Tdfc10 en Fibra DieteticaRosa Karen Suarez GuzmanNo ratings yet
- Sops For Quality Control in Small Size Breweries: Sandra HolmDocument50 pagesSops For Quality Control in Small Size Breweries: Sandra HolmjucalimaNo ratings yet
- Amylase AG XXL September 2003 Technical Present. CStu-1Document18 pagesAmylase AG XXL September 2003 Technical Present. CStu-1Diego Matias RamonNo ratings yet
- Chapter 2 NutritionDocument20 pagesChapter 2 NutritionAzie Nurul AkhtarNo ratings yet
- Executive Summary Red Rice WineDocument2 pagesExecutive Summary Red Rice WineJhoann CanenciaNo ratings yet
- Carbohydrate Chemistry Digestion and TransportDocument14 pagesCarbohydrate Chemistry Digestion and TransportManila MedNo ratings yet
- Biology Notes For IGCSEDocument54 pagesBiology Notes For IGCSErajeshn186% (36)
- CH 1-DigestionDocument127 pagesCH 1-Digestionmajdkaraki9No ratings yet
- Dgestion and Absorption of CHODocument15 pagesDgestion and Absorption of CHOessaNo ratings yet
- Effect of Anions on Amylase ActivityDocument10 pagesEffect of Anions on Amylase ActivityDaniel Alexander Black100% (1)
- Unit 6 Biology Genetic Modification & MicroorganismDocument7 pagesUnit 6 Biology Genetic Modification & MicroorganismWendyNo ratings yet
- High Fructose Syrup ProductionDocument4 pagesHigh Fructose Syrup ProductionPriyank LashkariNo ratings yet
- Food ChemistryDocument21 pagesFood Chemistrylinm@kilvington.vic.edu.auNo ratings yet
- Project On Natural PolymersDocument23 pagesProject On Natural PolymersChandraneel SinghNo ratings yet
- Enzyme Application in The Textile IndustryDocument14 pagesEnzyme Application in The Textile IndustryFie100% (1)
- Amylase EnzymeDocument6 pagesAmylase EnzymeDianne Faye ManabatNo ratings yet
- Amilase FB - 4004Document3 pagesAmilase FB - 4004Muhammad Aasim HassanNo ratings yet
- Explore Amylase Specificity Lab ReportDocument4 pagesExplore Amylase Specificity Lab ReportDevina Afraditya PavetaNo ratings yet
- Raw Materials Selection and Medium Development For Industrial Fermentation ProcessesDocument10 pagesRaw Materials Selection and Medium Development For Industrial Fermentation ProcessesIsaacNo ratings yet
- Enzymatic Hydrolysis of Wheat Starch For Glucose Syrup ProductionDocument10 pagesEnzymatic Hydrolysis of Wheat Starch For Glucose Syrup ProductionAndres Felipe Tovar BetancurNo ratings yet
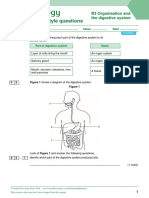
- Exam-Style Questions: B3 Organisation and The Digestive SystemDocument6 pagesExam-Style Questions: B3 Organisation and The Digestive Systemchemphycomb100% (1)
- Salivary Amylase ProjectDocument14 pagesSalivary Amylase ProjectTanmay SharmaNo ratings yet