You might also like
- Live Cell Assays: From Research to Regulatory ApplicationsFrom EverandLive Cell Assays: From Research to Regulatory ApplicationsNo ratings yet
- Autofluorescence - With - Supplementary Information-1Document27 pagesAutofluorescence - With - Supplementary Information-1kasa_10No ratings yet
- Cell Biology Assays: ProteinsFrom EverandCell Biology Assays: ProteinsFanny JaulinNo ratings yet
- Sensors 09 00378Document8 pagesSensors 09 00378gpaivNo ratings yet
- Cells 09 00838Document17 pagesCells 09 00838SueNo ratings yet
- C.L.I.P - Final VersionDocument9 pagesC.L.I.P - Final VersionJan KrajniakNo ratings yet
- Immunostaining of Voltage-Gated Ion Channels in Cell Lines and Neurons - Key Concepts and Potential PitfallsDocument27 pagesImmunostaining of Voltage-Gated Ion Channels in Cell Lines and Neurons - Key Concepts and Potential Pitfallskjf185No ratings yet
- Macrophage-Stimulating Peptides VKGFY and Cyclo (VKGFY) Act Through Nonopioid B-Endorphin ReceptorsDocument8 pagesMacrophage-Stimulating Peptides VKGFY and Cyclo (VKGFY) Act Through Nonopioid B-Endorphin Receptorsvanessa_murillo_5No ratings yet
- Miniaturizing Chemistry and Biology in MDocument16 pagesMiniaturizing Chemistry and Biology in MsuggestionboxNo ratings yet
- Detection of Nanometer-Sized Particles in Living Cells Using Modern Uorescence Uctuation MethodsDocument8 pagesDetection of Nanometer-Sized Particles in Living Cells Using Modern Uorescence Uctuation MethodsJaime JaimexNo ratings yet
- tmp992F TMPDocument10 pagestmp992F TMPFrontiersNo ratings yet
- 8 HalamansdbDocument14 pages8 HalamansdbbanglarkNo ratings yet
- Association Mucine NeuropepidesDocument4 pagesAssociation Mucine NeuropepidesAinaNo ratings yet
- Cerebrolysin Increases Recovery and Decreases Inflammation in An Autoimmune Model of Autoimmune EncephalitisDocument15 pagesCerebrolysin Increases Recovery and Decreases Inflammation in An Autoimmune Model of Autoimmune EncephalitisAlin MihaiNo ratings yet
- Research ArticleDocument12 pagesResearch ArticleKarthiNo ratings yet
- Ijo 06 06 744Document8 pagesIjo 06 06 744Ratih Kusuma DewiNo ratings yet
- PEG-Induced PCC in Human and Rodent CellsDocument15 pagesPEG-Induced PCC in Human and Rodent CellsembcelNo ratings yet
- Ozone Therapy Ameliorates Tubulointerstitial Inflammation by Regulating TLR4 in Adenine Induced CKD RatsDocument10 pagesOzone Therapy Ameliorates Tubulointerstitial Inflammation by Regulating TLR4 in Adenine Induced CKD RatsBoby FaesalNo ratings yet
- 1988 - Microencapsulation of Pancreatic Islet Cells - A Bioartificial Endocrine PancreasDocument6 pages1988 - Microencapsulation of Pancreatic Islet Cells - A Bioartificial Endocrine PancreasAndrés MenesesNo ratings yet
- Quantitative measurement of zinc secretion from pancreatic islets with high temporal resolutionDocument10 pagesQuantitative measurement of zinc secretion from pancreatic islets with high temporal resolutionAmirNo ratings yet
- Light-Induced Photon Emission by Mammalian CellsDocument5 pagesLight-Induced Photon Emission by Mammalian CellsAntonis TzambazakisNo ratings yet
- Determination of Reliable Biomass Indicators in The Cyanobacterium GloeotheceDocument9 pagesDetermination of Reliable Biomass Indicators in The Cyanobacterium GloeotheceInternational Journal of Chemical & Biological Sciences100% (2)
- Polymers: Noninvasive Tracking of mPEG-poly (Ala) Hydrogel-Embedded MIN6 Cells After Subcutaneous Transplantation in MiceDocument10 pagesPolymers: Noninvasive Tracking of mPEG-poly (Ala) Hydrogel-Embedded MIN6 Cells After Subcutaneous Transplantation in MiceMaialen Espinal ViguriNo ratings yet
- Cloning Expression Glucose Transport Protein: and Ofa MicrosomalDocument5 pagesCloning Expression Glucose Transport Protein: and Ofa MicrosomalSierra D. FranciscoNo ratings yet
- Sem4a Insulin1995 JBCDocument4 pagesSem4a Insulin1995 JBCAntonella Farabelli SuarezNo ratings yet
- Journal Pone 0085304Document13 pagesJournal Pone 0085304LollyCheveNo ratings yet
- Nanotoxicology Applied To Solid Lipid Nanoparticles and Nanostructured Lipid Carriers - A Systematic Review of in Vitro DataDocument18 pagesNanotoxicology Applied To Solid Lipid Nanoparticles and Nanostructured Lipid Carriers - A Systematic Review of in Vitro DataJonatas LopesNo ratings yet
- 1 s2.0 S0378517323001734 MainDocument14 pages1 s2.0 S0378517323001734 Mainnur handayaniNo ratings yet
- 3.evaluation of DFM, Isolation - IJPBS-July 2012Document5 pages3.evaluation of DFM, Isolation - IJPBS-July 2012SHIVA JYOTHINo ratings yet
- Gentamicin-Induced Apoptosis in Renal Cell Lines and Embryonic Rat FibroblastsDocument0 pagesGentamicin-Induced Apoptosis in Renal Cell Lines and Embryonic Rat FibroblastsCLPHtheoryNo ratings yet
- Physiology PharmacologyDocument126 pagesPhysiology PharmacologyuneedlesNo ratings yet
- Gene Therapy For Type 1 Diabetes Mellitus in Rats by Gastrointestinal Administration of Chitosan Nanoparticles Containing Human Insulin GeneDocument7 pagesGene Therapy For Type 1 Diabetes Mellitus in Rats by Gastrointestinal Administration of Chitosan Nanoparticles Containing Human Insulin GenesaifudinNo ratings yet
- Pharmaceutics 03 00497 PDFDocument13 pagesPharmaceutics 03 00497 PDFJai MurugeshNo ratings yet
- Articulo Laser 5Document6 pagesArticulo Laser 5oojonNo ratings yet
- 2 - Who Is in and Who Is OutDocument16 pages2 - Who Is in and Who Is OutMilorad ZjalićNo ratings yet
- Beclin 1Document8 pagesBeclin 1Arif MohiddinNo ratings yet
- Zhang - 2017 - Quantification of Lactate Isotopologues and IsotopomersDocument12 pagesZhang - 2017 - Quantification of Lactate Isotopologues and IsotopomersMokr Guerrero EsperanzaNo ratings yet
- FUT6 Promotes HCC Growth by Activating PI3K/Akt PathwayDocument7 pagesFUT6 Promotes HCC Growth by Activating PI3K/Akt PathwayArthur ClarkNo ratings yet
- News Letter For Industrial User Group: Summary of Project Background and ObjectivesDocument7 pagesNews Letter For Industrial User Group: Summary of Project Background and Objectivesaudrey_pogetNo ratings yet
- tmp3D5 TMPDocument9 pagestmp3D5 TMPFrontiersNo ratings yet
- tmp7286 TMPDocument15 pagestmp7286 TMPFrontiersNo ratings yet
- Koo Ken 2014Document7 pagesKoo Ken 2014Perla Andrea Urriola AraosNo ratings yet
- Violet Diode-Assisted Photoporation and Transfection of Cells - 2005Document7 pagesViolet Diode-Assisted Photoporation and Transfection of Cells - 2005Antonio Vázquez MotaNo ratings yet
- Jove Protocol 56627 Isolation Respiratory Measurements Mitochondria From ArabidopsisDocument10 pagesJove Protocol 56627 Isolation Respiratory Measurements Mitochondria From Arabidopsisfofo reminNo ratings yet
- A Multi-Chamber Microfluidic Intestinal Barrier Model Using Caco-2 Cells For Drug Transport StudiesDocument23 pagesA Multi-Chamber Microfluidic Intestinal Barrier Model Using Caco-2 Cells For Drug Transport StudiesGina PistolNo ratings yet
- Identifying Nuclear Mobility FactorsDocument13 pagesIdentifying Nuclear Mobility FactorsZhengwen ZhangNo ratings yet
- Journal Pcbi 1010708Document28 pagesJournal Pcbi 1010708hilaire0902No ratings yet
- Vitrification Preserves SSCS, Wyns 2013 PDFDocument12 pagesVitrification Preserves SSCS, Wyns 2013 PDFVarawenNo ratings yet
- Measurement of Glucose Homeostasis in VivoDocument10 pagesMeasurement of Glucose Homeostasis in Vivo19400668No ratings yet
- Exd 13489Document21 pagesExd 13489ShafiraNo ratings yet
- tmpD126 TMPDocument9 pagestmpD126 TMPFrontiersNo ratings yet
- Lab On A Chip: Miniaturisation For Chemistry, Physics, Biology, Materials Science and BioengineeringDocument9 pagesLab On A Chip: Miniaturisation For Chemistry, Physics, Biology, Materials Science and BioengineeringMadhu MidhaNo ratings yet
- ArticleDocument13 pagesArticlezulfikarNo ratings yet
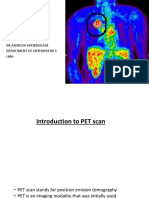
- Pet ScanDocument40 pagesPet ScanAkhilesh KhobragadeNo ratings yet
- Ricafort - Finals Journal Critiquing IS LectureDocument2 pagesRicafort - Finals Journal Critiquing IS Lecturesean ricafortNo ratings yet
- Cisplatin Cell Cicle ArrestDocument7 pagesCisplatin Cell Cicle ArrestGabriele KrauseNo ratings yet
- Evelyn Gabriela Gonzalez SimbañaDocument6 pagesEvelyn Gabriela Gonzalez SimbañaMOnzerratheManzzanitaSixxNo ratings yet
- JNR 2011 PDFDocument7 pagesJNR 2011 PDFPankaj RathoreNo ratings yet
- _Zinc Fixation for Flow Cytometry Analysis of Intracellular and Surface Epitopes, DNA Content, and Cell Proliferation_. In_ Current Protocols in CytometryDocument9 pages_Zinc Fixation for Flow Cytometry Analysis of Intracellular and Surface Epitopes, DNA Content, and Cell Proliferation_. In_ Current Protocols in CytometryPatricia GomesNo ratings yet
- Saloni Chaudhary 2019659564 Botony Assignment 2Document6 pagesSaloni Chaudhary 2019659564 Botony Assignment 2Saloni ChaudharyNo ratings yet
- Bab IiDocument1 pageBab IiBara Bagus RamandaNo ratings yet
- Bab IiDocument1 pageBab IiBara Bagus RamandaNo ratings yet
- Admin A 10 1 168 Cd5e504Document5 pagesAdmin A 10 1 168 Cd5e504Bara Bagus RamandaNo ratings yet
- Template Osce Station CtsDocument4 pagesTemplate Osce Station CtsBara Bagus RamandaNo ratings yet
- Herpetic Geometric Glossitis: Acyclovir Resistant Case in A Patient With Acute Myelogenous LeukemiaDocument3 pagesHerpetic Geometric Glossitis: Acyclovir Resistant Case in A Patient With Acute Myelogenous LeukemiaBara Bagus RamandaNo ratings yet
- Recurrent ITrichosporon Asahiii Glossitis A Case ReportDocument7 pagesRecurrent ITrichosporon Asahiii Glossitis A Case ReportBara Bagus RamandaNo ratings yet
- Tugas Jantung AnikDocument5 pagesTugas Jantung AnikBara Bagus RamandaNo ratings yet
- Admin A 10 1 168 Cd5e504Document5 pagesAdmin A 10 1 168 Cd5e504Bara Bagus RamandaNo ratings yet
- Recurrent ITrichosporon Asahiii Glossitis A Case ReportDocument7 pagesRecurrent ITrichosporon Asahiii Glossitis A Case ReportBara Bagus RamandaNo ratings yet
- Tugas Jantung AnikDocument5 pagesTugas Jantung AnikBara Bagus RamandaNo ratings yet
- Click To Edit Master Subtitle StyleDocument4 pagesClick To Edit Master Subtitle StyleBara Bagus RamandaNo ratings yet
- Daftar PustakaDocument2 pagesDaftar PustakaBara Bagus RamandaNo ratings yet
- 597 PDFDocument5 pages597 PDFBara Bagus RamandaNo ratings yet
- HumanEye PDFDocument16 pagesHumanEye PDFsneha Mary AntonyNo ratings yet
- Digestion and absorption multiple choice questionsDocument46 pagesDigestion and absorption multiple choice questionsAmbika dawadiNo ratings yet
- 9 Yoga Class Sequences for Different TargetsDocument20 pages9 Yoga Class Sequences for Different TargetsLaerteNo ratings yet
- TITLE The Effect of Carbon Dioxide Concentration on PhotosynthesisDocument5 pagesTITLE The Effect of Carbon Dioxide Concentration on PhotosynthesisDiana NurulNo ratings yet
- Unit - I Artificial Neural NetworksDocument23 pagesUnit - I Artificial Neural NetworksMary MorseNo ratings yet
- Neurohumoral Transmission - Meaning and Steps - AnimalsDocument9 pagesNeurohumoral Transmission - Meaning and Steps - AnimalsPragnesh ParmarNo ratings yet
- CommunionDocument6 pagesCommunionDavid Calvert100% (1)
- Exam ReviewDocument9 pagesExam ReviewjaxtradeNo ratings yet
- Pathfast PresepsinDocument6 pagesPathfast PresepsinWulan Ervinna SimanjuntakNo ratings yet
- 10k Training Program (Interval, Cruise and Long Dist)Document6 pages10k Training Program (Interval, Cruise and Long Dist)kevinalmerNo ratings yet
- 1.05 Psychobiology: Niels Birbaumer University of Tuèbingen, Germany and Herta Flor Humboldt-University, Berlin, GermanyDocument58 pages1.05 Psychobiology: Niels Birbaumer University of Tuèbingen, Germany and Herta Flor Humboldt-University, Berlin, GermanySandra Carolina Valencia LaraNo ratings yet
- Sinclaire AcademyDocument684 pagesSinclaire AcademyElijah Jane Stephanie Bernaldez93% (14)
- ds103 Evolution Vocabulary BingoDocument6 pagesds103 Evolution Vocabulary Bingoapi-110789702No ratings yet
- Vital Signs Taking - Procedures-Rationale-Checklist (New)Document8 pagesVital Signs Taking - Procedures-Rationale-Checklist (New)Davi ciiNo ratings yet
- Guide to Physical Assessment DocumentationDocument3 pagesGuide to Physical Assessment Documentationsandaman2225No ratings yet
- Transport in AnimalsDocument81 pagesTransport in AnimalsIbrahim NOORZADNo ratings yet
- Canine ReproductionDocument16 pagesCanine ReproductionDr rigs100% (2)
- Pathology For The Health Professions 5th Edition Damjanov Solutions ManualDocument25 pagesPathology For The Health Professions 5th Edition Damjanov Solutions ManualKevinRobertsbfak98% (56)
- Effect of Different Physical and Chemical Treatments On Detoxification of Ricin in Castor CakeDocument10 pagesEffect of Different Physical and Chemical Treatments On Detoxification of Ricin in Castor CakeL D Mendoza UrbinaNo ratings yet
- Range of MotionDocument17 pagesRange of MotionJim Cariaga100% (2)
- Stephanie M. Davis CVDocument5 pagesStephanie M. Davis CVStephanie Meyers DavisNo ratings yet
- Anaphy TransesDocument21 pagesAnaphy Transesfaith A.No ratings yet
- Labreport V SMSH 23 10141161 PDFDocument2 pagesLabreport V SMSH 23 10141161 PDFMohanNo ratings yet
- CHE 475 Biochemical Engineering II: Course SyllabusDocument19 pagesCHE 475 Biochemical Engineering II: Course SyllabusAkib Atique Khan DhruboNo ratings yet
- Visual Processing Disorder in Children OOJ 2 113Document3 pagesVisual Processing Disorder in Children OOJ 2 113Sharon ChristantoNo ratings yet
- ABG - Acid Base BalanceDocument4 pagesABG - Acid Base BalanceTingCheungNo ratings yet
- Novel Metabolic and Physiological Functions of Branched Chain Amino Acids: A ReviewDocument12 pagesNovel Metabolic and Physiological Functions of Branched Chain Amino Acids: A ReviewThiago Franco GuimarãesNo ratings yet
- Norvell ARG and Role in Renal InnervationDocument11 pagesNorvell ARG and Role in Renal InnervationGNo ratings yet
- Fisiologi Sistem Pencernaan: Kristanti, DRDocument64 pagesFisiologi Sistem Pencernaan: Kristanti, DRFeny Maharani100% (1)
- Silkworm moth: from egg to emergenceDocument78 pagesSilkworm moth: from egg to emergenceSoubhik MondalNo ratings yet
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Alex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessFrom EverandAlex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessNo ratings yet
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Periodic Tales: A Cultural History of the Elements, from Arsenic to ZincFrom EverandPeriodic Tales: A Cultural History of the Elements, from Arsenic to ZincRating: 3.5 out of 5 stars3.5/5 (137)
- Dark Matter and the Dinosaurs: The Astounding Interconnectedness of the UniverseFrom EverandDark Matter and the Dinosaurs: The Astounding Interconnectedness of the UniverseRating: 3.5 out of 5 stars3.5/5 (69)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- The Other End of the Leash: Why We Do What We Do Around DogsFrom EverandThe Other End of the Leash: Why We Do What We Do Around DogsRating: 5 out of 5 stars5/5 (63)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Water: The Epic Struggle for Wealth, Power, and CivilizationFrom EverandWater: The Epic Struggle for Wealth, Power, and CivilizationRating: 3.5 out of 5 stars3.5/5 (37)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- The Revolutionary Genius of Plants: A New Understanding of Plant Intelligence and BehaviorFrom EverandThe Revolutionary Genius of Plants: A New Understanding of Plant Intelligence and BehaviorRating: 4.5 out of 5 stars4.5/5 (137)