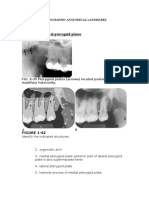
You might also like
- Creuzet 2005Document13 pagesCreuzet 2005asdNo ratings yet
- Hemichordate ArticleDocument8 pagesHemichordate ArticleCharu LathaNo ratings yet
- Evolution of The JawDocument13 pagesEvolution of The JawCraig MawsonNo ratings yet
- Acoel DevelopmentDocument6 pagesAcoel DevelopmentPaola BTNo ratings yet
- Review Article: Mohammad SarwarDocument17 pagesReview Article: Mohammad SarwarRuslan MilevskyNo ratings yet
- Developmental Evolution: This Side of Paradise: Anthony Graham and Imelda McgonnellDocument3 pagesDevelopmental Evolution: This Side of Paradise: Anthony Graham and Imelda McgonnellAroa ChansNo ratings yet
- Radiology IbqDocument48 pagesRadiology IbqSwati Pathak GiriNo ratings yet
- Bio 07 K2 (Kedah) AnsDocument13 pagesBio 07 K2 (Kedah) AnsFerguson TehNo ratings yet
- 9w HoloprosencephalyDocument4 pages9w HoloprosencephalyVũ Quang ĐăngNo ratings yet
- Derivation of The Mammalian Skull Vault: Gillianm - Morriss-KayDocument9 pagesDerivation of The Mammalian Skull Vault: Gillianm - Morriss-KaygandhiayuNo ratings yet
- 2 Head and Neck Anatomy SPTH 204Document79 pages2 Head and Neck Anatomy SPTH 204Noor KhalilNo ratings yet
- Kostka 1991Document12 pagesKostka 1991Lola BoaNo ratings yet
- Formery 2023 Molecular Evidence of AnteroposteriDocument20 pagesFormery 2023 Molecular Evidence of Anteroposterilei xuNo ratings yet
- Medial and Lateral Pterygoid Plates: Periapical Film A) MaxillaDocument25 pagesMedial and Lateral Pterygoid Plates: Periapical Film A) MaxillaPayal JainNo ratings yet
- Developmental Dynamics - 2008 - Hu - Effect of Bone Morphogenetic Protein Signaling On Development of The Jaw SkeletonDocument11 pagesDevelopmental Dynamics - 2008 - Hu - Effect of Bone Morphogenetic Protein Signaling On Development of The Jaw Skeletonhennysusanto18No ratings yet
- Evolution of The Embryonic DevelopmDocument12 pagesEvolution of The Embryonic Developmsatish kumarNo ratings yet
- Pharyngeal ApparatusDocument63 pagesPharyngeal Apparatusmobolajiadeaina7No ratings yet
- 2017 - HanEtAl - Deusterostomes Cambrian - LectureEcdysozoaDocument14 pages2017 - HanEtAl - Deusterostomes Cambrian - LectureEcdysozoaJavier PautaNo ratings yet
- Latam AnatomiaDocument11 pagesLatam AnatomiaMary MontoyaNo ratings yet
- Evolutionary Biology CohnDocument3 pagesEvolutionary Biology CohnCarlos Raul GomezNo ratings yet
- Disorders of Prosencephalic Development: ReviewDocument15 pagesDisorders of Prosencephalic Development: ReviewAna MariaNo ratings yet
- Tsukushi Functions As An Organizer Inducer by Inhibition of - 2004 - DevelopmenDocument12 pagesTsukushi Functions As An Organizer Inducer by Inhibition of - 2004 - Developmenhalil sahanNo ratings yet
- Pediatrics: The Journal ofDocument11 pagesPediatrics: The Journal ofRohini TondaNo ratings yet
- Tawk Et Al 2007 PDFDocument4 pagesTawk Et Al 2007 PDFCarliLoezarMartínezNo ratings yet
- Zollikofer Et Al - Evidence For Interpersonal Violence in The St. Césaire NeanderthalDocument5 pagesZollikofer Et Al - Evidence For Interpersonal Violence in The St. Césaire NeanderthalLuis Lobato de FariaNo ratings yet
- Embryology and Development of The Enteric Nervous System: H M Young, C J Hearn, D F NewgreenDocument3 pagesEmbryology and Development of The Enteric Nervous System: H M Young, C J Hearn, D F Newgreenproject-247758No ratings yet
- Unilateral CP PDFDocument29 pagesUnilateral CP PDFSuryaAtmajayaNo ratings yet
- Ni Hms 393042Document19 pagesNi Hms 393042Mutiara Tri JunisaNo ratings yet
- Development of The Juxta-Oral Organ in Rat EmbryoDocument7 pagesDevelopment of The Juxta-Oral Organ in Rat EmbryoMackarenna NarvaezNo ratings yet
- Yoshizawa 2014Document5 pagesYoshizawa 2014veqmahwxqzzimuqymoNo ratings yet
- C - Takeuchi, Nature, 1999, LimbDocument7 pagesC - Takeuchi, Nature, 1999, LimbplNo ratings yet
- Cranial Morphology in The Brachygnathic SheepDocument9 pagesCranial Morphology in The Brachygnathic SheepguadialvarezNo ratings yet
- Specialized Craniofacial Anatomy of A Titanosaurian Embryo From ArgentinaDocument19 pagesSpecialized Craniofacial Anatomy of A Titanosaurian Embryo From ArgentinaJorge BarNo ratings yet
- Apidima Cave Fossils Provide EarliestDocument21 pagesApidima Cave Fossils Provide Earliestevripidis tziokasNo ratings yet
- Origen Humano 2019Document21 pagesOrigen Humano 2019Fausto CésarNo ratings yet
- HHS Public Access: On The Morphogenesis of FeathersDocument12 pagesHHS Public Access: On The Morphogenesis of FeathersLinda FebrianiNo ratings yet
- Poll 3 ZoologyDocument2 pagesPoll 3 ZoologysurajmeherakawfdNo ratings yet
- 1 s2.0 S0960982214003145 MainDocument5 pages1 s2.0 S0960982214003145 MainGmod GopnikNo ratings yet
- PCR-Based Identification of Culex Pipiens Complex Collected in JapanDocument8 pagesPCR-Based Identification of Culex Pipiens Complex Collected in JapanSohan kunduNo ratings yet
- Endoscopic Ethmoidectomy - FESS - Surgical TechniqueDocument29 pagesEndoscopic Ethmoidectomy - FESS - Surgical TechniquekityamuwesiNo ratings yet
- Baume - Ontogenesis AtmDocument13 pagesBaume - Ontogenesis AtmDANIELA NARVAEZ GUAÑARITANo ratings yet
- Adc 8810Document5 pagesAdc 8810dna1979No ratings yet
- Anatomie Os Mandibulaire J.ydbio.2004.08.046Document12 pagesAnatomie Os Mandibulaire J.ydbio.2004.08.046Free CoursesNo ratings yet
- Michael J. F. Barresi, Scott F. Gilbert - Developmental Biology (2018, Sinauer Associates) (Dragged)Document3 pagesMichael J. F. Barresi, Scott F. Gilbert - Developmental Biology (2018, Sinauer Associates) (Dragged)Mackarenna NarvaezNo ratings yet
- Hagfish (Cyclostomata, Vertebrata) : Searching For The Ancestral Developmental Plan of VertebratesDocument6 pagesHagfish (Cyclostomata, Vertebrata) : Searching For The Ancestral Developmental Plan of VertebratesJ.D. NobleNo ratings yet
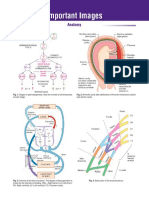
- Important ImagesDocument37 pagesImportant ImagesSubhani MahapatraNo ratings yet
- Topohistology of Sympathetic and ParasymDocument11 pagesTopohistology of Sympathetic and ParasymOfe CosNo ratings yet
- Case Report: A Rare Stapes AbnormalityDocument4 pagesCase Report: A Rare Stapes AbnormalityCarolina UrrutiaNo ratings yet
- 2019 Evolution of The Chordate TelencephalonDocument16 pages2019 Evolution of The Chordate TelencephalonFabian ArenasNo ratings yet
- 1 s2.0 S0965174822000807 MainDocument9 pages1 s2.0 S0965174822000807 MainRogelio FloresNo ratings yet
- Restoring Auditory Cortex Plasticity in Adult Mice by Restricting Thalamic Adenosine SignalingDocument6 pagesRestoring Auditory Cortex Plasticity in Adult Mice by Restricting Thalamic Adenosine SignalingErikSmithNo ratings yet
- Somites Without A Clock 2014 Dias Et AlDocument9 pagesSomites Without A Clock 2014 Dias Et AlCarmen CoelhoNo ratings yet
- Cranial Base ScottDocument30 pagesCranial Base ScottShirley AliagaNo ratings yet
- Balemans 2007Document9 pagesBalemans 2007malliasarahannNo ratings yet
- Identification Guide To Antarctic CopepodsDocument42 pagesIdentification Guide To Antarctic CopepodsmauricioNo ratings yet
- Nuclear Pore ComplexesDocument9 pagesNuclear Pore ComplexesMNo ratings yet
- Bacilus Coagulans SporesDocument13 pagesBacilus Coagulans SporesscribdNo ratings yet
- Elaeis Guineensis 7Document6 pagesElaeis Guineensis 7Jesus Llorente mendozaNo ratings yet
- Elaeis Guineensis 6Document14 pagesElaeis Guineensis 6Jesus Llorente mendozaNo ratings yet
- Elaeis Guineensis 8Document9 pagesElaeis Guineensis 8Jesus Llorente mendozaNo ratings yet
- Enterolobium Cyclocarpum 3Document21 pagesEnterolobium Cyclocarpum 3Jesus Llorente mendozaNo ratings yet
- Sapium Glandulosum 2Document7 pagesSapium Glandulosum 2Jesus Llorente mendozaNo ratings yet
- Almuqarrabun 2014Document12 pagesAlmuqarrabun 2014serena morenaNo ratings yet
- Samanea Saman 8Document7 pagesSamanea Saman 8Jesus Llorente mendozaNo ratings yet
- Albizia Saman 1Document20 pagesAlbizia Saman 1Jesus Llorente mendozaNo ratings yet
- 20951-Article Text-64675-1-10-20180406Document7 pages20951-Article Text-64675-1-10-20180406Rosalie CarganilloNo ratings yet
- Samanea Saman 6Document6 pagesSamanea Saman 6Jesus Llorente mendozaNo ratings yet
- Terminalia Catappa 12Document10 pagesTerminalia Catappa 12Jesus Llorente mendozaNo ratings yet
- Acacia Mangium 4Document3 pagesAcacia Mangium 4Jesus Llorente mendozaNo ratings yet
- Acacia Mangium 5Document10 pagesAcacia Mangium 5Jesus Llorente mendozaNo ratings yet
- Acacia Mangium 3Document16 pagesAcacia Mangium 3Jesus Llorente mendozaNo ratings yet
- Acacia Mangium 2Document12 pagesAcacia Mangium 2Jesus Llorente mendozaNo ratings yet
- 2175 FullDocument12 pages2175 FullJesus Llorente mendozaNo ratings yet
- RibozymesDocument29 pagesRibozymesJesus Llorente mendozaNo ratings yet
- 2525 FullDocument10 pages2525 FullJesus Llorente mendozaNo ratings yet
- Pharyngeal Arch Patterning in The Absence of Neural CrestDocument4 pagesPharyngeal Arch Patterning in The Absence of Neural CrestJesus Llorente mendozaNo ratings yet
- Applied Psychology 29 8 18Document52 pagesApplied Psychology 29 8 18SoouNo ratings yet
- Neuromarketing - Applications, Challenges and Promises - BiomedresDocument14 pagesNeuromarketing - Applications, Challenges and Promises - BiomedresRitika RituNo ratings yet
- ADHDDocument21 pagesADHDYuniita VerayantiiNo ratings yet
- NSSBIO3E PPT Ch15 eDocument336 pagesNSSBIO3E PPT Ch15 eLau Wing YanNo ratings yet
- The Neurological ExaminationDocument53 pagesThe Neurological ExaminationApolinar González Hernández100% (2)
- A. Sound FoundationDocument47 pagesA. Sound FoundationKashmira TadaNo ratings yet
- DR - Roezwir SP.SDocument76 pagesDR - Roezwir SP.SSyifaAsriFauziahNo ratings yet
- Polygraphy Definition of TermsDocument14 pagesPolygraphy Definition of TermsMark Ranggas80% (20)
- Manav Techreport1Document27 pagesManav Techreport1Sehaj SrkNo ratings yet
- Neurobiology of AddictionDocument57 pagesNeurobiology of AddictionJef_8No ratings yet
- Comparison of UMN & LMN SyndromesDocument2 pagesComparison of UMN & LMN SyndromesPJHGNo ratings yet
- CXC CSEC HSB January 2017 P2 PDFDocument17 pagesCXC CSEC HSB January 2017 P2 PDFSayyid Muhammad Aqeed AabidiNo ratings yet
- El Insomnio en La Infancia PDFDocument10 pagesEl Insomnio en La Infancia PDFCarlos Perez DiazNo ratings yet
- Cingulate EpilepsyDocument18 pagesCingulate EpilepsyfeawefNo ratings yet
- Histology of Nervous SystemDocument59 pagesHistology of Nervous SystemMelissa Aina Mohd YusofNo ratings yet
- Acoustic NeuromaDocument23 pagesAcoustic Neuromacefiroth100% (1)
- Ebp Cedera Kepala - The Effect of Giving Oxygenation With Simple Oxygen Mask andDocument6 pagesEbp Cedera Kepala - The Effect of Giving Oxygenation With Simple Oxygen Mask andNindy kusuma wardaniNo ratings yet
- Multiple Choice QuizDocument7 pagesMultiple Choice QuizNargess OsmanNo ratings yet
- Taste (Gustation)Document29 pagesTaste (Gustation)kityamuwesi100% (1)
- Ophthalmology Set 4Document6 pagesOphthalmology Set 4ajay khadeNo ratings yet
- Concerns About Infrasound From Wind TurbinesDocument10 pagesConcerns About Infrasound From Wind TurbinesgeofflevNo ratings yet
- InterLaminar Window DescriptionDocument6 pagesInterLaminar Window DescriptionTiger PowerNo ratings yet
- 2002 - Spiking Neuron Models - Single Neurons, Populations, Plasticity PDFDocument564 pages2002 - Spiking Neuron Models - Single Neurons, Populations, Plasticity PDFSebastianRamirezNo ratings yet
- Neurology 2012 MrcppassDocument154 pagesNeurology 2012 MrcppassRaouf Ra'fat SolimanNo ratings yet
- Cybernetic Organism 30092013125043 Cybernetic OrganismDocument32 pagesCybernetic Organism 30092013125043 Cybernetic OrganismSânthú Stày'sNo ratings yet
- HNNE Con Signos de AlarmaDocument1 pageHNNE Con Signos de AlarmayocondaariasNo ratings yet
- Liceo de Cagayan University: College of Medical Laboratory ScienceDocument3 pagesLiceo de Cagayan University: College of Medical Laboratory ScienceLol lolNo ratings yet
- Exp Physiol 2011 Cahill 528 38Document11 pagesExp Physiol 2011 Cahill 528 38Cristopher Castro RdNo ratings yet
- Toyota Kata: Mobilizing Our Ingenuity Through Good ManagementDocument26 pagesToyota Kata: Mobilizing Our Ingenuity Through Good ManagementVali100% (1)
- Piriformis Syndrome: A Narrative Review of The Anatomy, Diagnosis, and TreatmentDocument10 pagesPiriformis Syndrome: A Narrative Review of The Anatomy, Diagnosis, and TreatmentHumairah AnandaNo ratings yet