You might also like
- TNF Skews Monocyte Di Erentiation From Macrophages To Dendritic CellsDocument9 pagesTNF Skews Monocyte Di Erentiation From Macrophages To Dendritic CellsmfcostaNo ratings yet
- Rheumatology-2014-Liu-rheumatology-keu316 HormonaDocument11 pagesRheumatology-2014-Liu-rheumatology-keu316 HormonaBluna0291No ratings yet
- Dex3 and Survivin-2B: Two Novel Splice Variants of The ApoptosisDocument6 pagesDex3 and Survivin-2B: Two Novel Splice Variants of The ApoptosisNona NonicaaNo ratings yet
- Investigation of A Combination of Amiodarone and Itraconazole For Treatment of American Trypanosomiasis in Dogs Madigan Et Al 2019Document13 pagesInvestigation of A Combination of Amiodarone and Itraconazole For Treatment of American Trypanosomiasis in Dogs Madigan Et Al 2019Maritza NunezNo ratings yet
- Trypanosomes PDFDocument15 pagesTrypanosomes PDFMarcela RomeroNo ratings yet
- COVID-19 Makes B Cells Forget, But T Cells Remember: PreviewsDocument3 pagesCOVID-19 Makes B Cells Forget, But T Cells Remember: PreviewsRamy Refaat LewisNo ratings yet
- Test 30Document15 pagesTest 30CiprianNo ratings yet
- Tian 2016Document13 pagesTian 2016Epiphany SonderNo ratings yet
- FB2008A0024Document5 pagesFB2008A0024Giga Hasabi AlkaraniNo ratings yet
- Pentoxifylline Reduces Inflammation and Prevents Myocardial Perfusion Derangements in Experimental Chronic Chagas' CardiomyopathyDocument11 pagesPentoxifylline Reduces Inflammation and Prevents Myocardial Perfusion Derangements in Experimental Chronic Chagas' CardiomyopathyDenise MayumiNo ratings yet
- Acido ValproicoDocument10 pagesAcido ValproicopreedtooneNo ratings yet
- Acidosalicilico ExossomaDocument7 pagesAcidosalicilico ExossomaRenato GranadoNo ratings yet
- Acute Opisthorchiasis in A Kazakh ManDocument2 pagesAcute Opisthorchiasis in A Kazakh Manhusna aninda farahitaNo ratings yet
- 566 Letters: A Unifying Hypothesis Is That Activation of Na+/K+Document2 pages566 Letters: A Unifying Hypothesis Is That Activation of Na+/K+saadsffggeettrgbfbfbftggrg ergrtertererefrerrNo ratings yet
- Oncogenic Function of TUSC3 in Non-Small Cell Lung Cancer Is AssociatedDocument12 pagesOncogenic Function of TUSC3 in Non-Small Cell Lung Cancer Is AssociatedMárcia TrindadeNo ratings yet
- Correos Electrónicos 5688Document11 pagesCorreos Electrónicos 5688Lorena ChaparroNo ratings yet
- Zeng 2016Document7 pagesZeng 2016Thanasis PapatheodorouNo ratings yet
- T-Helper (1) Cytokines Increase During Early Pregnancy in Women With A History of Recurrent Spontaneous AbortionDocument5 pagesT-Helper (1) Cytokines Increase During Early Pregnancy in Women With A History of Recurrent Spontaneous AbortionChu Chu gaNo ratings yet
- Poster Presentations Experimental Hematology 44 (2016) S56-S110Document1 pagePoster Presentations Experimental Hematology 44 (2016) S56-S110aymenNo ratings yet
- CD4 CD25 Foxp3 Regulatory T Cells in Myelodysplastic Syndrome (MDS)Document5 pagesCD4 CD25 Foxp3 Regulatory T Cells in Myelodysplastic Syndrome (MDS)María Alejandra Lopez SalasNo ratings yet
- ER Stress-Related Molecules Induced by Hantaan Virus InfectionDocument10 pagesER Stress-Related Molecules Induced by Hantaan Virus InfectionJulia MouraNo ratings yet
- Antuñano 2000 PDFDocument10 pagesAntuñano 2000 PDFAlonso FlNo ratings yet
- Expression of A Breast-Cancer-Associated Protein (Ps2) in Human Neuro-Endocrine TumoursDocument5 pagesExpression of A Breast-Cancer-Associated Protein (Ps2) in Human Neuro-Endocrine TumoursSarly FebrianaNo ratings yet
- Treatment of Human Disease by Adeno-Associated Viral Gene TransferDocument33 pagesTreatment of Human Disease by Adeno-Associated Viral Gene Transfergauri ThapaNo ratings yet
- Apoptosis and Cancer: Cckchao@mail - Cgu.edu - TWDocument16 pagesApoptosis and Cancer: Cckchao@mail - Cgu.edu - TWsari1No ratings yet
- Nature 11647Document5 pagesNature 11647Jef_8No ratings yet
- 2001 Nador VirologyDocument9 pages2001 Nador VirologyleandrosarvanitakisNo ratings yet
- CRM1 Inhibition Promotes Cytotoxicity in Ewing Sarcoma Cells by Repressing EWS-FLI1 - Dependent IGF-1 Signaling 1Document12 pagesCRM1 Inhibition Promotes Cytotoxicity in Ewing Sarcoma Cells by Repressing EWS-FLI1 - Dependent IGF-1 Signaling 1janetchen1996125No ratings yet
- Journal Pbio 3000850Document33 pagesJournal Pbio 3000850Juan David Pardo Mr. PostmanNo ratings yet
- 1 s2.0 S0021915021003269 MainDocument1 page1 s2.0 S0021915021003269 MainIoanna NikdimaNo ratings yet
- Butlertjaden 2013Document15 pagesButlertjaden 2013Raluca MosoiaNo ratings yet
- Chagas Therapy NEJM 2011Document8 pagesChagas Therapy NEJM 2011Ever LuizagaNo ratings yet
- 15 JTH 12782Document12 pages15 JTH 12782Ricardo GomezNo ratings yet
- Insertional Mutagenesis Identifies Multiple Networks of Cooperating Genes Driving Intestinal TumorigenesisDocument10 pagesInsertional Mutagenesis Identifies Multiple Networks of Cooperating Genes Driving Intestinal Tumorigenesistalha saleemNo ratings yet
- 2271Document5 pages2271Diana CsillaNo ratings yet
- Tvmp50 Is An Immunogenic Metalloproteinase During Male TrichomoniasisDocument12 pagesTvmp50 Is An Immunogenic Metalloproteinase During Male Trichomoniasiscarolramos71No ratings yet
- Hong, 2011Document9 pagesHong, 2011Dana SchneiderNo ratings yet
- Tmp77a1 TMPDocument14 pagesTmp77a1 TMPFrontiersNo ratings yet
- Possible Mechanisms of Disease Development in Tuberous SclerosisDocument7 pagesPossible Mechanisms of Disease Development in Tuberous Sclerosisplastioid4079No ratings yet
- Tenascin-C Promotes Tumor Cell Migration and Metastasis Through Integrin A9b1-Mediated YAP InhibitionDocument12 pagesTenascin-C Promotes Tumor Cell Migration and Metastasis Through Integrin A9b1-Mediated YAP InhibitionSayda DhaouadiNo ratings yet
- Ni Hms 72445Document16 pagesNi Hms 72445Bianca BujderNo ratings yet
- 48497487development of A Real-Time PCR Assay For20160901-31954-5rom1y-Libre - pdf147277084Document6 pages48497487development of A Real-Time PCR Assay For20160901-31954-5rom1y-Libre - pdf147277084milixza botacioNo ratings yet
- Leptin 1Document9 pagesLeptin 1vestibuleNo ratings yet
- The Sympathetic Nervous System Modulates CD4+FoxP3+ Regulatory T Cells Via A TGFDocument9 pagesThe Sympathetic Nervous System Modulates CD4+FoxP3+ Regulatory T Cells Via A TGFPedro FonsecaNo ratings yet
- Chronic Inflammation in Fat Plays A Crucial Role in The Development of Obesity-Related Insulin ResistanceDocument10 pagesChronic Inflammation in Fat Plays A Crucial Role in The Development of Obesity-Related Insulin ResistanceRachel Lalaine Marie SialanaNo ratings yet
- Expression of A Mutant Form of Leishmania Donovani Centrin Reduces The Growth of The ParasiteDocument10 pagesExpression of A Mutant Form of Leishmania Donovani Centrin Reduces The Growth of The ParasitemclimacoNo ratings yet
- Nifuroxazide Inhibits Survival of Multiple Myeloma Cells by Directly Inhibiting STAT3Document9 pagesNifuroxazide Inhibits Survival of Multiple Myeloma Cells by Directly Inhibiting STAT3Nella Rahayu SusaniNo ratings yet
- Presentación 27Document18 pagesPresentación 27Felipe RayoNo ratings yet
- TTDocument15 pagesTTHusni FaridNo ratings yet
- Expression of Fas (CD95:APO-1) and Fas Ligand (FasL) in Experimentally-Induced Acute PancreatitisDocument9 pagesExpression of Fas (CD95:APO-1) and Fas Ligand (FasL) in Experimentally-Induced Acute PancreatitismattpauziNo ratings yet
- Articulo 1 InmunogenomicaDocument6 pagesArticulo 1 InmunogenomicaMichelle PoulainNo ratings yet
- The Pathophysiology of Pure Red Cell Aplasia: Implications For TherapyDocument9 pagesThe Pathophysiology of Pure Red Cell Aplasia: Implications For TherapyPhương NhungNo ratings yet
- tmp120F TMPDocument10 pagestmp120F TMPFrontiersNo ratings yet
- Pancreatic TumorDocument11 pagesPancreatic TumorPatricia BezneaNo ratings yet
- Celecoxib Combined With Salirasib Strongly Inhibits Pancreatic Cancer Cells in 2D and 3D CulturesDocument8 pagesCelecoxib Combined With Salirasib Strongly Inhibits Pancreatic Cancer Cells in 2D and 3D CulturesRoyNo ratings yet
- Autoimmunity in Chagas' DiseaseDocument4 pagesAutoimmunity in Chagas' DiseaseValNo ratings yet
- J. Biol. Chem.-1996-Couet-4545-52Document9 pagesJ. Biol. Chem.-1996-Couet-4545-52eze033No ratings yet
- Epitope Mapping of Trans-Sialidase From Trypanosoma Cruzi RevealsDocument7 pagesEpitope Mapping of Trans-Sialidase From Trypanosoma Cruzi RevealsmclimacoNo ratings yet
- Association of Mast CellsDocument8 pagesAssociation of Mast CellsJéssica MacêdoNo ratings yet
- Anders Et Al., 2014Document22 pagesAnders Et Al., 2014Tatiana Sanchez AlvarezNo ratings yet
- Arneth Et Al., 2017 Insulina Humana TratamientoDocument8 pagesArneth Et Al., 2017 Insulina Humana TratamientoTatiana Sanchez AlvarezNo ratings yet
- Almarzooqi Et Al., 2018 LDMDocument8 pagesAlmarzooqi Et Al., 2018 LDMTatiana Sanchez AlvarezNo ratings yet
- Ali-Boucetta2011 MTTDocument14 pagesAli-Boucetta2011 MTTTatiana Sanchez AlvarezNo ratings yet
- Alya Et Al., 2011Document18 pagesAlya Et Al., 2011Tatiana Sanchez AlvarezNo ratings yet
- Ali Et Al-2016-Cancer ScienceDocument8 pagesAli Et Al-2016-Cancer ScienceTatiana Sanchez AlvarezNo ratings yet
- Rapid Deployment of A Mobile Biosafety Level-3 LabDocument15 pagesRapid Deployment of A Mobile Biosafety Level-3 LabTatiana Sanchez AlvarezNo ratings yet
- Axon Glycoprotein Routing in Nerve Polarity, Function, and RepairDocument12 pagesAxon Glycoprotein Routing in Nerve Polarity, Function, and RepairTatiana Sanchez AlvarezNo ratings yet
- GKO v2 User ManualDocument15 pagesGKO v2 User ManualTatiana Sanchez AlvarezNo ratings yet
- DAPI Nucleic Acid Stain: Material Amount Storage StabilityDocument5 pagesDAPI Nucleic Acid Stain: Material Amount Storage StabilityTatiana Sanchez AlvarezNo ratings yet
- Mitosox™ Red Mitochondrial Superoxide Indicator For Live-Cell Imaging (M36008)Document3 pagesMitosox™ Red Mitochondrial Superoxide Indicator For Live-Cell Imaging (M36008)Tatiana Sanchez AlvarezNo ratings yet
- Generation of Growth CurveDocument1 pageGeneration of Growth CurveTatiana Sanchez AlvarezNo ratings yet
- Gotu Kola (Centella Asiatica)Document11 pagesGotu Kola (Centella Asiatica)My Pham Thi DiemNo ratings yet
- Analysis Physalis Angulata L - After ProofreadingDocument7 pagesAnalysis Physalis Angulata L - After ProofreadingPT BUTTA MAMMINASATA SHARIANo ratings yet
- Effect of Oncoxin Oral Solution in HER2-Overexpressing Breast CancerDocument12 pagesEffect of Oncoxin Oral Solution in HER2-Overexpressing Breast Cancerleonaldy sintesahealthNo ratings yet
- Intl Journal of Cancer - 2014 - Lakritz - Beneficial Bacteria Stimulate Host Immune Cells To Counteract Dietary and GeneticDocument12 pagesIntl Journal of Cancer - 2014 - Lakritz - Beneficial Bacteria Stimulate Host Immune Cells To Counteract Dietary and GeneticTrang MốcNo ratings yet
- Modelling Avascular Tumour Growth: Helen M. ByrneDocument46 pagesModelling Avascular Tumour Growth: Helen M. ByrnederNo ratings yet
- Inflammation and Neoplasia ApoptosisDocument6 pagesInflammation and Neoplasia ApoptosisTapitNo ratings yet
- Rmes 000734Document8 pagesRmes 000734Javier MartinNo ratings yet
- Referat HIEDocument14 pagesReferat HIErifkipspdNo ratings yet
- ProlpDocument25 pagesProlpherryNo ratings yet
- 3865-3871 2.23 Aliye Aras (MINI-REVIEW)Document7 pages3865-3871 2.23 Aliye Aras (MINI-REVIEW)Marcela FernandesNo ratings yet
- EJCHEM Volume 65 Issue 8 Pages 43-55Document13 pagesEJCHEM Volume 65 Issue 8 Pages 43-55فردوس يسNo ratings yet
- Muricata Fruit: Anticancer Activity of Acetogenins From AnnonaDocument10 pagesMuricata Fruit: Anticancer Activity of Acetogenins From AnnonananaNo ratings yet
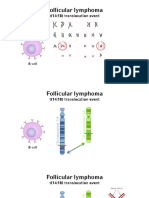
- Follicular Lymphoma: T (14:18) Translocation EventDocument15 pagesFollicular Lymphoma: T (14:18) Translocation EventAntoaneta PapNo ratings yet
- The Anti Cancer Essential Oil ReferenceDocument9 pagesThe Anti Cancer Essential Oil ReferenceΡαφαέλα ΠηλείδηNo ratings yet
- MVMJ - Volume 23 - Issue 2 - Pages 31-39Document9 pagesMVMJ - Volume 23 - Issue 2 - Pages 31-39HebaNo ratings yet
- The Effects of Zhikang Capsule On The Growth and Apoptosis of Breast Cancer Mcf7Document4 pagesThe Effects of Zhikang Capsule On The Growth and Apoptosis of Breast Cancer Mcf7Riya CassendraNo ratings yet
- Large Granular Lymphocytic Leukemia. Current Diagnostic and Therapeutic Approaches and Novel Treatment OptionsDocument29 pagesLarge Granular Lymphocytic Leukemia. Current Diagnostic and Therapeutic Approaches and Novel Treatment OptionsOrxan HumbetovNo ratings yet
- Neem, Pesticide and Medicinal ApplicationDocument60 pagesNeem, Pesticide and Medicinal Applicationlok8No ratings yet
- NanoparticlesDocument29 pagesNanoparticlesMOHAMMAD ARSHAD PATHANNo ratings yet
- Function of MitochondriaDocument5 pagesFunction of MitochondriaArshia NazirNo ratings yet
- Chapter 4 Mechanisms of Anticancer DrugsDocument13 pagesChapter 4 Mechanisms of Anticancer DrugsLarisa StoianNo ratings yet
- Pappas' Pysiology of The Cell ManualDocument66 pagesPappas' Pysiology of The Cell ManualVeres AndrásNo ratings yet
- Shilajit A Panacea For CancerDocument9 pagesShilajit A Panacea For CancerRakeshKumarNo ratings yet
- Lemon GrassDocument6 pagesLemon GrassKavisa GhoshNo ratings yet
- Pathoma - Chapter 1Document13 pagesPathoma - Chapter 1Jonathan AiresNo ratings yet
- Pathology Mcqs CorrectDocument45 pagesPathology Mcqs CorrectAhmed Alamin83% (6)
- Hoffbrands Haematology Online MCQSDocument29 pagesHoffbrands Haematology Online MCQSATHUMANI JUMBENo ratings yet
- Complement System Part II: Role in ImmunityDocument26 pagesComplement System Part II: Role in ImmunityAnca LunguNo ratings yet
- Role of Oxidative Stress in Semen Quality of Infertile MenDocument9 pagesRole of Oxidative Stress in Semen Quality of Infertile MenAyadPalaniNo ratings yet
- Male Subfertility and Oxidative StressssssssssssssssssssssDocument17 pagesMale Subfertility and Oxidative StressssssssssssssssssssssjonoNo ratings yet