You might also like
- MS-MLPA Protocol Two-Tube TMSP-V001Document17 pagesMS-MLPA Protocol Two-Tube TMSP-V001Felipe Ruiz BoteroNo ratings yet
- Lab 1.1 Protocol - Easy Lithium Acetate Transformation of Yeast - UPDATEDDocument10 pagesLab 1.1 Protocol - Easy Lithium Acetate Transformation of Yeast - UPDATEDcolby mcintoshNo ratings yet
- chemistry-technical-document-CHTD 500 v1 Revo 07jul2016 PDFDocument27 pageschemistry-technical-document-CHTD 500 v1 Revo 07jul2016 PDFMoni Becerra WongNo ratings yet
- chemistry-technical-document-CHTD 500 v1 revAA 07jul2016-AnyDocument42 pageschemistry-technical-document-CHTD 500 v1 revAA 07jul2016-AnyErikk DangNo ratings yet
- Numbering and Mapping of Genes On The ChromosomesDocument13 pagesNumbering and Mapping of Genes On The ChromosomesSreekuttyNo ratings yet
- Lecture Notes Algorithms in Bioinformatics I - Prof. Daniel HusonDocument28 pagesLecture Notes Algorithms in Bioinformatics I - Prof. Daniel HusonmsNo ratings yet
- Library Construction: Transcriptome MrnaDocument5 pagesLibrary Construction: Transcriptome MrnaDeepak MehtaNo ratings yet
- 2nd Bio AssgnmntDocument6 pages2nd Bio AssgnmntNitinmanubanshNo ratings yet
- ArtikelDocument1 pageArtikelfitraNo ratings yet
- Gene Expression Central DogmaDocument7 pagesGene Expression Central DogmaSualeha FatimaNo ratings yet
- Lecture 4 - 15 End Replication and Genetic Code - Transcription PDFDocument78 pagesLecture 4 - 15 End Replication and Genetic Code - Transcription PDFFafa MusNo ratings yet
- Worksheet Q3 Week 4&5 PDFDocument4 pagesWorksheet Q3 Week 4&5 PDFJaybie TejadaNo ratings yet
- CMMB-Fund Mol BioDocument45 pagesCMMB-Fund Mol BioMahmud RakibNo ratings yet
- Anatomy of A GeneDocument33 pagesAnatomy of A GenemskikiNo ratings yet
- Mrs. Ofelia Solano Saludar: Department of Natural Sciences University of St. La Salle Bacolod CityDocument33 pagesMrs. Ofelia Solano Saludar: Department of Natural Sciences University of St. La Salle Bacolod CitymskikiNo ratings yet
- Central Dogma FlowDocument8 pagesCentral Dogma FlowAvirup RayNo ratings yet
- DNA, RNA, Protein-Gene ExpressionDocument18 pagesDNA, RNA, Protein-Gene ExpressionFahim AbidNo ratings yet
- Gene to Protein: Transcription and Translation ExplainedDocument18 pagesGene to Protein: Transcription and Translation ExplainedMa. Joy JocosolNo ratings yet
- Bacterial Genetics in 40 CharactersDocument14 pagesBacterial Genetics in 40 Charactersعلي الكوافيNo ratings yet
- Non-coding DNA Functions and FractionsDocument5 pagesNon-coding DNA Functions and FractionsNikki SStarkNo ratings yet
- Genomes, Transcription, RegulationDocument8 pagesGenomes, Transcription, RegulationAngelica SmithNo ratings yet
- DNA, Genes, Chromosomes and Genomes ExplainedDocument41 pagesDNA, Genes, Chromosomes and Genomes ExplainedfirstylesNo ratings yet
- BCH 323 NotesDocument41 pagesBCH 323 NotesvictorNo ratings yet
- DNA and Chromosome Structure ExplainedDocument27 pagesDNA and Chromosome Structure ExplainedHumera SarwarNo ratings yet
- Genome Matt Ridley NotesDocument7 pagesGenome Matt Ridley Notesambiz2005No ratings yet
- Worksheet Q3 Week 4Document4 pagesWorksheet Q3 Week 4Jaybie TejadaNo ratings yet
- Nucleic Acid Structure and FunctionDocument37 pagesNucleic Acid Structure and FunctionLiz VillarNo ratings yet
- Microbial GeneticsDocument17 pagesMicrobial Geneticsزين العابدين محمد عويش مشريNo ratings yet
- Information Transfer: Central Dogma of Molecular BiologyDocument22 pagesInformation Transfer: Central Dogma of Molecular BiologyAvirup RayNo ratings yet
- 3.1 GenesDocument75 pages3.1 GenesBriguitte Subilete HuamaniNo ratings yet
- C Value ParadoxDocument4 pagesC Value ParadoxEr Purushottam PalNo ratings yet
- The Trypanosoma Cruzi Genome Project: Nuclear Karyotype and Gene Mapping of Clone CL BrenerDocument8 pagesThe Trypanosoma Cruzi Genome Project: Nuclear Karyotype and Gene Mapping of Clone CL BrenerSergio Guillermo Cuevas CarbonellNo ratings yet
- WbhffkndnckdcbudfjlnvskjcjancjabvswDocument4 pagesWbhffkndnckdcbudfjlnvskjcjancjabvswthirumalaivasan GNo ratings yet
- Central Dogma of Molecular BiologyDocument5 pagesCentral Dogma of Molecular BiologyHanumat Singh100% (1)
- RetroviruseDocument8 pagesRetroviruseMalik RamzanNo ratings yet
- Chapter On TranscriptomicsDocument13 pagesChapter On TranscriptomicsMazhar FarNo ratings yet
- Cytogenetics SemiFinal L TRANSESDocument15 pagesCytogenetics SemiFinal L TRANSESQUEEN NICOLE O. MACALAMNo ratings yet
- The Central Dogma of Molecular Biology Was First Stated by in 1958 and Re-Stated in A Paper Published in 1970Document3 pagesThe Central Dogma of Molecular Biology Was First Stated by in 1958 and Re-Stated in A Paper Published in 1970Mark Anthony LorenzoNo ratings yet
- InheritanceDocument9 pagesInheritanceFalak FathimaNo ratings yet
- Genome OrganisationDocument9 pagesGenome Organisationw5waNo ratings yet
- Chromosome StructureDocument11 pagesChromosome StructureAshley Nicole DelizoNo ratings yet
- The Biochemistry of DNA Replication and RepairDocument290 pagesThe Biochemistry of DNA Replication and RepairHas MasNo ratings yet
- Bacterial Genetics: DR - Tetty Aman Nasution M Med SCDocument59 pagesBacterial Genetics: DR - Tetty Aman Nasution M Med SCAndry SipahutarNo ratings yet
- Genetic Aspects of Rheumatic Diseases: Benedicte A. Lie, Marthe T. Mæhlen, Javier MartinDocument40 pagesGenetic Aspects of Rheumatic Diseases: Benedicte A. Lie, Marthe T. Mæhlen, Javier MartinAbuzdea AlexNo ratings yet
- DNA Kinetics ReassociationDocument4 pagesDNA Kinetics ReassociationSarmistha NayakNo ratings yet
- Band Methods 2Document7 pagesBand Methods 2Jazlynn WongNo ratings yet
- DNA and Chromosomes - 2019 PDFDocument89 pagesDNA and Chromosomes - 2019 PDFRiya PremNo ratings yet
- Block II Lecture 1: Recombinant DNA TechnologyDocument42 pagesBlock II Lecture 1: Recombinant DNA TechnologyRitesh Chandrakant SuryawanshiNo ratings yet
- Miniature Protein BIologyDocument83 pagesMiniature Protein BIologyPREET KANOOGANo ratings yet
- Biochem 14 Chap 24Document60 pagesBiochem 14 Chap 24'chaca' Dian Catur PermatasariNo ratings yet
- Genome MappingDocument9 pagesGenome Mappingrag.1607No ratings yet
- Biomed 08Document14 pagesBiomed 08api-3706483No ratings yet
- Principles of Microbiology 1 (Summary)Document8 pagesPrinciples of Microbiology 1 (Summary)Mabelle DucusinNo ratings yet
- Introduction To The Human Genome and Chromosome Structure: Prathap SDocument12 pagesIntroduction To The Human Genome and Chromosome Structure: Prathap Sprathap somuNo ratings yet
- Chapter IV Introduction To Bacterial Genetics-1Document67 pagesChapter IV Introduction To Bacterial Genetics-1temesgensemahegn55No ratings yet
- Chapter Summary: Chapter 4: The Structure and Function of GenesDocument26 pagesChapter Summary: Chapter 4: The Structure and Function of GenesmongguNo ratings yet
- Reverse Transcriptase (RT) Is An: WikipediaDocument4 pagesReverse Transcriptase (RT) Is An: Wikipedianmp274No ratings yet
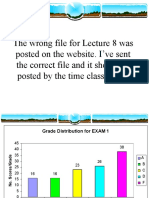
- The Wrong File For Lecture 8 Was Posted On The Website. I've Sent The Correct File and It Should Be Posted by The Time Class Is OutDocument51 pagesThe Wrong File For Lecture 8 Was Posted On The Website. I've Sent The Correct File and It Should Be Posted by The Time Class Is Outmbhattacharya094956No ratings yet
- Genetics Minimals FinalDocument5 pagesGenetics Minimals FinalМөнхгэрэл ГанбатNo ratings yet
- DNA Structure and Function GuideDocument15 pagesDNA Structure and Function GuideDoctor Muhammad JunaidNo ratings yet
- Human Chromosomes: An Illustrated Introduction to Human CytogeneticsFrom EverandHuman Chromosomes: An Illustrated Introduction to Human CytogeneticsRating: 5 out of 5 stars5/5 (1)
- Bio-Rad Explorer Cloning and Sequencing Explorer Series: Curriculum ManualDocument332 pagesBio-Rad Explorer Cloning and Sequencing Explorer Series: Curriculum ManualRNo ratings yet
- DNA Cloning: A 4-Step ProcessDocument26 pagesDNA Cloning: A 4-Step ProcessmadhuriNo ratings yet
- Sure Cut Buffers RocheDocument3 pagesSure Cut Buffers RochecvisnisNo ratings yet
- 4c. Full ProceedingDocument626 pages4c. Full ProceedingRato Petani Bandeng SilamonNo ratings yet
- Plasmid Cloning by Restriction Enzyme DigestDocument3 pagesPlasmid Cloning by Restriction Enzyme DigestVempati Rahul KumarNo ratings yet
- MAN0012966 CloneJET PCR Cloning 40rxn UG PDFDocument2 pagesMAN0012966 CloneJET PCR Cloning 40rxn UG PDFPriya JainNo ratings yet
- Production of Glutamate and Stereospecific Flavors, (S) - Linalool and (+) - Valencene, by Synechocystis Sp. PCC6803Document7 pagesProduction of Glutamate and Stereospecific Flavors, (S) - Linalool and (+) - Valencene, by Synechocystis Sp. PCC6803Fernando IzquierdoNo ratings yet
- LigasesDocument7 pagesLigasesSuraj PatilNo ratings yet
- Universal GenomeWalker 2.0 User Manual - 040314Document24 pagesUniversal GenomeWalker 2.0 User Manual - 040314xprakashNo ratings yet
- cDNA Synthesis System: Instruction ManualDocument36 pagescDNA Synthesis System: Instruction ManualmaniiiiiiiiNo ratings yet
- 03Q PDFDocument2 pages03Q PDFFCiênciasNo ratings yet
- Rapid and Efficient Cosmid Cloning: D.Ish-Horowicz and J.F.BurkeDocument10 pagesRapid and Efficient Cosmid Cloning: D.Ish-Horowicz and J.F.BurkerezqNo ratings yet
- Overview - DNA Cloning (Article) 2 - Khan AcademyDocument13 pagesOverview - DNA Cloning (Article) 2 - Khan AcademyMaryem SafdarNo ratings yet
- pgl3 Luciferase Reporter Vectors Protocol PDFDocument31 pagespgl3 Luciferase Reporter Vectors Protocol PDFSr. RZNo ratings yet
- pGEM-T EasyDocument15 pagespGEM-T EasyRocío Romero HinojosaNo ratings yet
- Lab ReportDocument7 pagesLab ReportAlliedschool DefencecampusNo ratings yet
- Analytical Techniques in BiotechnologyDocument25 pagesAnalytical Techniques in Biotechnologybs19b006 Ansh Ruhela100% (1)
- Why Johnny Can't Clone: Common Pitfalls and Not So Common SolutionsDocument10 pagesWhy Johnny Can't Clone: Common Pitfalls and Not So Common SolutionsGanesh KumarNo ratings yet
- Genomic Dna by Ligation SQK lsk110 GDE - 9108 - v110 - Revx - 10nov2020 MinionDocument26 pagesGenomic Dna by Ligation SQK lsk110 GDE - 9108 - v110 - Revx - 10nov2020 MinionOscar Evrard ArruéNo ratings yet
- Genetic Engineering Laboratory Manual: Prepared by Lilly M Saleena Department of Bioinformatics SRM UniversityDocument21 pagesGenetic Engineering Laboratory Manual: Prepared by Lilly M Saleena Department of Bioinformatics SRM UniversityShreeya BhatNo ratings yet
- CloneJET PCR Cloning KitDocument9 pagesCloneJET PCR Cloning Kitodile1987No ratings yet
- Dna Ligase, Terminal Transferase, Adapter and LinkerDocument80 pagesDna Ligase, Terminal Transferase, Adapter and LinkerSujit ShandilyaNo ratings yet
- Restriction EnzymeDocument28 pagesRestriction Enzymeanon_173640290No ratings yet
- User Guide: Targetron Gene Knockout SystemDocument28 pagesUser Guide: Targetron Gene Knockout SystemLuisNo ratings yet
- Recombinant DNA Technology (Siddra Ijaz, Imran Ul Haq) (Z-Library)Document157 pagesRecombinant DNA Technology (Siddra Ijaz, Imran Ul Haq) (Z-Library)Vera MenezesNo ratings yet
- Laboratory Manual For Genetic EngineeringDocument133 pagesLaboratory Manual For Genetic EngineeringRajesh Patel100% (2)