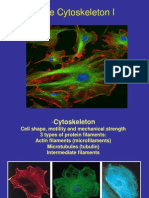
You might also like
- CytoskeletonDocument65 pagesCytoskeletonSapana SubediNo ratings yet
- Cytoskeleton Motor Proteins and ECM - SCDocument77 pagesCytoskeleton Motor Proteins and ECM - SCAlice RizzoNo ratings yet
- The Cytoskeleton and Cell MotilityDocument25 pagesThe Cytoskeleton and Cell MotilityRenz L. SalumbreNo ratings yet
- Cytoskeleton PRESENTATIONDocument36 pagesCytoskeleton PRESENTATIONChaudryNomiNo ratings yet
- DJ (Microtubules, Microfilaments and Intermediate Filaments) - DJGFDocument43 pagesDJ (Microtubules, Microfilaments and Intermediate Filaments) - DJGFNick_989893No ratings yet
- 1 7 CytoskeletonDocument31 pages1 7 CytoskeletonAditya AgrawalNo ratings yet
- 5 - Cytoskeleton and Cell MotilityDocument21 pages5 - Cytoskeleton and Cell Motilitywhether913No ratings yet
- MicrotubulesDocument39 pagesMicrotubulesAdnan QureshiNo ratings yet
- From Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.From EverandFrom Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.No ratings yet
- Cytoskeleton and Cell Motility 1Document40 pagesCytoskeleton and Cell Motility 1Muhamad Farhan RLNo ratings yet
- Lecture 6 - CytoskeletonDocument38 pagesLecture 6 - CytoskeletonNL DearestNo ratings yet
- L 4 Cell Organization IIDocument35 pagesL 4 Cell Organization IIsNo ratings yet
- C & M Bio CytoskeletonDocument40 pagesC & M Bio CytoskeletonA PNo ratings yet
- Lectures 8-10 NotesDocument15 pagesLectures 8-10 Notesapi-3721438No ratings yet
- The CytoskeletonDocument31 pagesThe CytoskeletonSanthoshsv 143No ratings yet
- Prepared By:-: Priyanka Yadav M.Sc. Life Sciences Ist SemDocument37 pagesPrepared By:-: Priyanka Yadav M.Sc. Life Sciences Ist SemleartaNo ratings yet
- Organization and Functions of Cell Cytoskeleton ComponentsDocument28 pagesOrganization and Functions of Cell Cytoskeleton ComponentsRNo ratings yet
- Biol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationDocument49 pagesBiol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationHui Ka HoNo ratings yet
- Cytoskeleton Structures for Cell Shape and OrganizationDocument49 pagesCytoskeleton Structures for Cell Shape and OrganizationHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationDocument49 pagesBiol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationHui Ka HoNo ratings yet
- The cytoskeletonDocument10 pagesThe cytoskeletonAhmed khanNo ratings yet
- Cytoskeleton - GROUP 2Document19 pagesCytoskeleton - GROUP 2n42fjjy8g7No ratings yet
- The Cytoskeleton Is A Network of Filament and Tubules That Extends Throughout A CellDocument9 pagesThe Cytoskeleton Is A Network of Filament and Tubules That Extends Throughout A CellHania GulNo ratings yet
- CELL CYTOSKELETONDocument7 pagesCELL CYTOSKELETONandersonmack047No ratings yet
- BM 1 1 CytoskeletonDocument24 pagesBM 1 1 CytoskeletonSanthoshi Sadhanaa SankarNo ratings yet
- Weeks 1 2 Cytoskeleton 2024Document97 pagesWeeks 1 2 Cytoskeleton 2024cavadzadenaibeNo ratings yet
- Biomolecules & Cells Biol10002 Lectures 1-9 Cell Biology Geoff Mcfadden, RM 211 Biosciences 2 (Old Botany)Document41 pagesBiomolecules & Cells Biol10002 Lectures 1-9 Cell Biology Geoff Mcfadden, RM 211 Biosciences 2 (Old Botany)waimoeNo ratings yet
- Cytoskeleton DADocument35 pagesCytoskeleton DAAnonymous NVBCqKWHNo ratings yet
- Microtubule Oranizing CenterDocument5 pagesMicrotubule Oranizing CenterSasha GutierrezNo ratings yet
- (B) Microfilaments: FunctionDocument2 pages(B) Microfilaments: FunctionDrbee10No ratings yet
- Thecytoskeleton PPTDocument11 pagesThecytoskeleton PPTseenu mohapatraNo ratings yet
- 2 Cytoskeleton & Cell MotilityDocument47 pages2 Cytoskeleton & Cell MotilityChristineNo ratings yet
- Enumerating The Structure of Major Protein, Describe The Eukaryotic CytoskeletonDocument10 pagesEnumerating The Structure of Major Protein, Describe The Eukaryotic CytoskeletonTathagata ChakravarttyNo ratings yet
- Cell Cycle Dependent Regulation of Microtubule Dynamics by Microtubules Associated Proteins (MAPs) and Its Misregulation Causes AneuploidyDocument4 pagesCell Cycle Dependent Regulation of Microtubule Dynamics by Microtubules Associated Proteins (MAPs) and Its Misregulation Causes AneuploidyEditor IJTSRDNo ratings yet
- Molecular Motors Nanobiotech FinalDocument24 pagesMolecular Motors Nanobiotech FinalHAIMANTI NATHNo ratings yet
- Molecular Bio Lecture 13Document38 pagesMolecular Bio Lecture 13AngryNo ratings yet
- He Cell 1Document34 pagesHe Cell 1Le Phuong LyNo ratings yet
- lecture 6 - cytoskeleton STDocument18 pageslecture 6 - cytoskeleton STminhph.23bi14469No ratings yet
- Microtubules - The Cell - NCBI BookshelfDocument5 pagesMicrotubules - The Cell - NCBI Bookshelfanulorance98No ratings yet
- L 2 Cell Structure and FunctionDocument25 pagesL 2 Cell Structure and Functionkaukab azimNo ratings yet
- Cytoplasm and InclusionDocument21 pagesCytoplasm and Inclusionelphas walelaNo ratings yet
- AMINO ACIDS AND PROTEINS - THE CELL'S CYTOSKELETON AND ITS FUNCTIONS (38 charactersDocument2 pagesAMINO ACIDS AND PROTEINS - THE CELL'S CYTOSKELETON AND ITS FUNCTIONS (38 charactersyejiNo ratings yet
- The Cytoskeleton I: Actin, Microtubules and Intermediate FilamentsDocument62 pagesThe Cytoskeleton I: Actin, Microtubules and Intermediate FilamentsRebecca ReillyNo ratings yet
- The CytoskeletonDocument9 pagesThe CytoskeletonElena OlmedoNo ratings yet
- CHAP 2. CSH TẾ BÀO - 232Document100 pagesCHAP 2. CSH TẾ BÀO - 232Nguyễn Anh QuangNo ratings yet
- 11.sitoskeleton Dan Pergerakan Sel-Dr - IsraDocument40 pages11.sitoskeleton Dan Pergerakan Sel-Dr - IsraTa RaNo ratings yet
- Cytoskeleton 13.4.20Document39 pagesCytoskeleton 13.4.20NikhilNo ratings yet
- Pi Is 0960982206017714Document2 pagesPi Is 0960982206017714AjengFadhillahNo ratings yet
- CytosceletonDocument41 pagesCytosceletonaidar.seralinNo ratings yet
- Microtubules - The CytoskeletonDocument11 pagesMicrotubules - The Cytoskeletonhafssa BachirNo ratings yet
- Lecture 3 - Chapter 8-Cytoskeleton ADocument75 pagesLecture 3 - Chapter 8-Cytoskeleton AKw Chan33% (3)
- Cellular MovementsDocument27 pagesCellular MovementsngidizzmNo ratings yet
- Understanding the Dynamic CytoskeletonDocument66 pagesUnderstanding the Dynamic CytoskeletonChristian Santiago100% (1)
- Mb4-Cytoskeleton and Intercellular MatrixDocument40 pagesMb4-Cytoskeleton and Intercellular Matrixfaezeh zare karizakNo ratings yet
- Cytoskeleton Molecular Motors: Structures and Their Functions in NeuronDocument10 pagesCytoskeleton Molecular Motors: Structures and Their Functions in NeuronnadaNo ratings yet
- Cytoskeleton roles in cell motility and structureDocument2 pagesCytoskeleton roles in cell motility and structureDavid YapNo ratings yet
- Babs1201 Notes On CytoskeletonDocument1 pageBabs1201 Notes On CytoskeletonMatthew WongNo ratings yet
- Bio 49 Activity 1Document6 pagesBio 49 Activity 1Theresa LambayonNo ratings yet
- Histology Cytoplasm Part 4 Lecture - Cytoskeleton and Intermediate FilamentsDocument5 pagesHistology Cytoplasm Part 4 Lecture - Cytoskeleton and Intermediate FilamentsKahlen SivanNo ratings yet
- Cy To SkeletonDocument17 pagesCy To SkeletonKitkat AlorroNo ratings yet
- 1-Metabolism Introduction-E7-1187Document14 pages1-Metabolism Introduction-E7-1187nathanNo ratings yet
- MBB 321 Tutorial 1 PDFDocument4 pagesMBB 321 Tutorial 1 PDFnathanNo ratings yet
- 6 Polysaccharides E7 1181 PDFDocument36 pages6 Polysaccharides E7 1181 PDFnathanNo ratings yet
- BPK 205 Intro To Human Physiology: Lecture 12 - Sensory Physiology I: Intro & The Somatic SensesDocument24 pagesBPK 205 Intro To Human Physiology: Lecture 12 - Sensory Physiology I: Intro & The Somatic SensesnathanNo ratings yet
- AAMC Content Outline General Math and Scientific Inquiry SkillsDocument165 pagesAAMC Content Outline General Math and Scientific Inquiry SkillsadfhNo ratings yet
- Chapter 4Document29 pagesChapter 4skywalkerNo ratings yet
- Lectures 8-10 NotesDocument15 pagesLectures 8-10 Notesapi-3721438No ratings yet
- Disorders of Lysosome-Related Organelle BiogenesisDocument34 pagesDisorders of Lysosome-Related Organelle BiogenesisMaria AnggieNo ratings yet
- (Advances in Biochemical Engineering - Biotechnology 119) Takayuki Nishizaka (Auth.), Isao Endo, Teruyuki Nagamune (Eds.) - Nano - Micro Biotechnology (2010, Springer-Verlag Berlin Heidelberg) PDFDocument271 pages(Advances in Biochemical Engineering - Biotechnology 119) Takayuki Nishizaka (Auth.), Isao Endo, Teruyuki Nagamune (Eds.) - Nano - Micro Biotechnology (2010, Springer-Verlag Berlin Heidelberg) PDFAlejandra EscalonaNo ratings yet
- Cytoskeleton 13.4.20Document39 pagesCytoskeleton 13.4.20NikhilNo ratings yet
- CytoskeletonDocument86 pagesCytoskeletonAbid Al RezaNo ratings yet
- Molecular Biology of The Cell, Sixth Edition Chapter 16: The CytoskeletonDocument37 pagesMolecular Biology of The Cell, Sixth Edition Chapter 16: The CytoskeletonJeanPaule Joumaa100% (1)
- Molecular Bio Lecture 13Document38 pagesMolecular Bio Lecture 13AngryNo ratings yet
- Test Bank For Biology How Life Works Second EditionDocument33 pagesTest Bank For Biology How Life Works Second EditionJanetSmithcwdz100% (36)
- Cell Structure and FunctionDocument44 pagesCell Structure and FunctionPedro SuyuNo ratings yet
- Cytoskeleton Molecular Motors: Structures and Their Functions in NeuronDocument10 pagesCytoskeleton Molecular Motors: Structures and Their Functions in NeuronnadaNo ratings yet
- Chapter 3: Compartmentation: Cells and Tissues: Functional Compartments of The BodyDocument10 pagesChapter 3: Compartmentation: Cells and Tissues: Functional Compartments of The BodyShivani RailowalNo ratings yet
- (Lewis I. Held JR) Quirks of Human Anatomy An Ev PDFDocument273 pages(Lewis I. Held JR) Quirks of Human Anatomy An Ev PDFNancy Morales100% (2)
- Biological Science Canadian 2Nd Edition Freeman Solutions Manual Full Chapter PDFDocument38 pagesBiological Science Canadian 2Nd Edition Freeman Solutions Manual Full Chapter PDFkieranthang03m100% (14)
- UT Dallas Syllabus For Biol5440.001.09s Taught by Gail Breen (Breen)Document5 pagesUT Dallas Syllabus For Biol5440.001.09s Taught by Gail Breen (Breen)UT Dallas Provost's Technology GroupNo ratings yet
- MBB231 Chapter 9 Cytoskeleton Introduction 1197Document28 pagesMBB231 Chapter 9 Cytoskeleton Introduction 1197nathanNo ratings yet
- Molecular MotorDocument21 pagesMolecular MotorKetan PatelNo ratings yet
- (Microbiologia) Modern Food Microbiology 6th Ed - James M. JDocument9 pages(Microbiologia) Modern Food Microbiology 6th Ed - James M. JDewiakyuNo ratings yet
- Bio MotorsDocument16 pagesBio MotorsNeha Srivastava100% (2)
- 2 Cytoskeleton & Cell MotilityDocument47 pages2 Cytoskeleton & Cell MotilityChristineNo ratings yet
- Chapter 6 Ap Bio NotesDocument16 pagesChapter 6 Ap Bio NotescatieevansNo ratings yet
- Molecular MotorsDocument13 pagesMolecular MotorsEzri ChilengweNo ratings yet
- Chapter 17 - Cell Organization and Movement I: MicrofilamentsDocument64 pagesChapter 17 - Cell Organization and Movement I: MicrofilamentsRebecca Long HeiseNo ratings yet
- AP Biology Name - Chapter 6 Guided Reading AssignmentDocument6 pagesAP Biology Name - Chapter 6 Guided Reading AssignmentRobert Ladaga33% (3)
- 01 02 Cytoskeleton PDFDocument12 pages01 02 Cytoskeleton PDFsigninNo ratings yet
- Introduction To Molecular Motor Proteins: Part 1 Lecture NotesDocument12 pagesIntroduction To Molecular Motor Proteins: Part 1 Lecture Noteskinza888No ratings yet
- Bio 150 Trans 3.1Document13 pagesBio 150 Trans 3.1Jolaine ValloNo ratings yet
- Physio EssenceDocument602 pagesPhysio EssencePhonogaming XNo ratings yet