You might also like
- The Archaeology of Large-Scale Manipulation of Prey: The Economic and Social Dynamics of Mass HuntingFrom EverandThe Archaeology of Large-Scale Manipulation of Prey: The Economic and Social Dynamics of Mass HuntingKristen A. CarlsonNo ratings yet
- New Fission Processes in The Zoanthid Palythoa Caribaeorum Description and Quantitative AspectsDocument26 pagesNew Fission Processes in The Zoanthid Palythoa Caribaeorum Description and Quantitative AspectsAlejandro GarciaNo ratings yet
- Evidence of Gonochorism in A Grouper, Mycteroperca Rosacea, From The Gulf of California, MexicoDocument12 pagesEvidence of Gonochorism in A Grouper, Mycteroperca Rosacea, From The Gulf of California, MexicocristianNo ratings yet
- Amariles2017 - Article - FoodResourcePartitioningOfTheM ART M.lunuLATUS Y HENLEI de NAVIA ColombiaDocument16 pagesAmariles2017 - Article - FoodResourcePartitioningOfTheM ART M.lunuLATUS Y HENLEI de NAVIA ColombiaPerla RosalesNo ratings yet
- Cheng2020 Article Genome-wideSNPsRevealComplexFiDocument15 pagesCheng2020 Article Genome-wideSNPsRevealComplexFinabilaaa yasmin hakimNo ratings yet
- Hernaezand Wehrtmann 2007 Populationbiologyof CallichirusseilacheriDocument13 pagesHernaezand Wehrtmann 2007 Populationbiologyof Callichirusseilacherikeltony aquinoNo ratings yet
- Mytilus Eduli M. TrossulusDocument13 pagesMytilus Eduli M. TrossulusSuany Quesada CalderonNo ratings yet
- Tripp Valdez 2010Document6 pagesTripp Valdez 2010Cecilia Oñate BustosNo ratings yet
- Population Genetics and Phylogeography of Sea TurtlesDocument23 pagesPopulation Genetics and Phylogeography of Sea Turtleshunterdog42No ratings yet
- Hippo CampusDocument12 pagesHippo CampusDevinder KumarNo ratings yet
- Revealing Stock-Recruitment Relationships in Lobsters and Crabs: Is Experimental Ecology The Key?Document30 pagesRevealing Stock-Recruitment Relationships in Lobsters and Crabs: Is Experimental Ecology The Key?DanielaNo ratings yet
- Ajb 90 6 915 PDFDocument9 pagesAjb 90 6 915 PDFsalma sabilaNo ratings yet
- 2 PDFDocument13 pages2 PDFVianey Sánchez FigueroaNo ratings yet
- Peces Amazonia Germination-SeedsDocument12 pagesPeces Amazonia Germination-SeedsEmilio CamposantoNo ratings yet
- Journal of Phycology - 2019 - Fraser - The Biogeographic Importance of Buoyancy in Macroalgae A Case Study of The SouthernDocument14 pagesJournal of Phycology - 2019 - Fraser - The Biogeographic Importance of Buoyancy in Macroalgae A Case Study of The SouthernSebastian CavourNo ratings yet
- Dermochelys Coriacea SkeletonDocument13 pagesDermochelys Coriacea SkeletonYessica VargasNo ratings yet
- The Role of The Andean Uplift As An Asymmetrical BDocument22 pagesThe Role of The Andean Uplift As An Asymmetrical BValeriaNo ratings yet
- Graham-Et Al, 2010, Leatherback & JellyfishDocument10 pagesGraham-Et Al, 2010, Leatherback & JellyfishJonas CorreaNo ratings yet
- Predation and The Abundance of Juvenile Blue Crabs: A Comparison of Selected East and Gulf Coast (Usa) StudiesDocument7 pagesPredation and The Abundance of Juvenile Blue Crabs: A Comparison of Selected East and Gulf Coast (Usa) StudiesTomás Damián Mora PodioNo ratings yet
- Takahashi and Checkeley 2008 (OK)Document8 pagesTakahashi and Checkeley 2008 (OK)Ivan De Jesus MendozaNo ratings yet
- Fouquet 2007Document16 pagesFouquet 2007Carol AraújoNo ratings yet
- A Collective Navigation Hypothesis For Homeward Migration in Anadromous SalmonidsDocument18 pagesA Collective Navigation Hypothesis For Homeward Migration in Anadromous SalmonidsÁngelGarcíaDiestroNo ratings yet
- J. Shellfish Res 2008 C. Corteziensis Growth Gametogenic Activity CIDocument11 pagesJ. Shellfish Res 2008 C. Corteziensis Growth Gametogenic Activity CIRomina AlterioNo ratings yet
- Palumbi 2006 The History of Whales Read From DNADocument14 pagesPalumbi 2006 The History of Whales Read From DNAEliza ZanahNo ratings yet
- Residency and Abundance of Sperm Whales (Physeter Macrocephalus) in Nemuro Strait, Hokkaido, JapanDocument11 pagesResidency and Abundance of Sperm Whales (Physeter Macrocephalus) in Nemuro Strait, Hokkaido, JapanGianni PavanNo ratings yet
- Parra2009 PDFDocument9 pagesParra2009 PDFJorge Luis SantamariaNo ratings yet
- Global Phylogeography of The Loggerhead Turtle (Caretta Caretta) As Indicated by Mitochondrial Dna Haplotypes4Document9 pagesGlobal Phylogeography of The Loggerhead Turtle (Caretta Caretta) As Indicated by Mitochondrial Dna Haplotypes4Takada NamaNo ratings yet
- Pejhanmehr 2022Document170 pagesPejhanmehr 2022georgesidneybarachoNo ratings yet
- Endemic Fishes of The Gulf of CaliforniaDocument12 pagesEndemic Fishes of The Gulf of CaliforniaMarianne MtzNo ratings yet
- Costa-Souza Et Al 2019 - Growth, Sexual Maturity and Dimorphism in Six Species of SnappingDocument14 pagesCosta-Souza Et Al 2019 - Growth, Sexual Maturity and Dimorphism in Six Species of Snappingkeltony aquinoNo ratings yet
- Missouri Botanical Garden Press Annals of The Missouri Botanical GardenDocument10 pagesMissouri Botanical Garden Press Annals of The Missouri Botanical GardenFrancione GomesNo ratings yet
- Beckett Et Al 2007Document18 pagesBeckett Et Al 2007Alan BermudezNo ratings yet
- Restrepo Et Al. 2022. Evaluation of Putative Hybrid Hatchlings Between Hawksbill and Green Sea Turtles in Tortuguero, Costa RicaDocument8 pagesRestrepo Et Al. 2022. Evaluation of Putative Hybrid Hatchlings Between Hawksbill and Green Sea Turtles in Tortuguero, Costa RicaIrama PerozoNo ratings yet
- Asteraceae - Cuello de Botella - 2000 PDFDocument8 pagesAsteraceae - Cuello de Botella - 2000 PDFEsteban FVNo ratings yet
- Population Genetics of Four Heavily Exploited Shark Species Around The Arabian PeninsulaDocument16 pagesPopulation Genetics of Four Heavily Exploited Shark Species Around The Arabian Peninsulaaljazza03No ratings yet
- Evolution of Habitat Use by Deep-Sea Mussels: Marine Biology February 2006Document12 pagesEvolution of Habitat Use by Deep-Sea Mussels: Marine Biology February 2006joeNo ratings yet
- Koch 2006 Molecules and Migration BiogeographDocument22 pagesKoch 2006 Molecules and Migration BiogeographsalarNo ratings yet
- Global Ecology and Conservation: Original Research ArticleDocument13 pagesGlobal Ecology and Conservation: Original Research ArticleKatie CrolleyNo ratings yet
- Calis Et Al., 2005Document14 pagesCalis Et Al., 2005Sergio LlorenteNo ratings yet
- Sphyrna Lewini: Global Phylogeography of The Scalloped Hammerhead SharkDocument13 pagesSphyrna Lewini: Global Phylogeography of The Scalloped Hammerhead SharkRodrigo MazzoleniNo ratings yet
- Dispersal and Life History of Brown Widow Spiders in Dated Invasive Populations On Two ContinentsDocument11 pagesDispersal and Life History of Brown Widow Spiders in Dated Invasive Populations On Two Continentsairtonschubert014No ratings yet
- Elephant SealDocument9 pagesElephant SealAbraham MartinezNo ratings yet
- tmp4033 TMPDocument5 pagestmp4033 TMPFrontiersNo ratings yet
- Age-At-Death Standards For Mesoamerican Prehispanic and Colonial Infant, Child, and Juvenile SkeletonsDocument18 pagesAge-At-Death Standards For Mesoamerican Prehispanic and Colonial Infant, Child, and Juvenile SkeletonsLuisa Fernanda GonzalezNo ratings yet
- Molluscan Studies: Journal ofDocument7 pagesMolluscan Studies: Journal ofJAVIER ANDRES VENEGAS OJEDANo ratings yet
- Is A New Paradigm Emerging For Oceanic Island Biogeography?: Guest EditorialDocument5 pagesIs A New Paradigm Emerging For Oceanic Island Biogeography?: Guest EditorialT.J. KnotekNo ratings yet
- Bloom 2012Document15 pagesBloom 2012Fadhli LatuconsinaNo ratings yet
- Phylogeography of The Chilean Red Cricket Cratomelus Cryptic Diversity in Central ChileDocument16 pagesPhylogeography of The Chilean Red Cricket Cratomelus Cryptic Diversity in Central ChileJoaquín Eduardo Sepúlveda AstudilloNo ratings yet
- Sex Ratios of Loggerhead Sea Turtles Caretta Caretta During The Juvenile Pelagic StageDocument13 pagesSex Ratios of Loggerhead Sea Turtles Caretta Caretta During The Juvenile Pelagic StagealejoolduNo ratings yet
- Undersea Constellations: The Global Biology of An Endangered Marine Megavertebrate Further Informed Through Citizen ScienceDocument15 pagesUndersea Constellations: The Global Biology of An Endangered Marine Megavertebrate Further Informed Through Citizen SciencekrisNo ratings yet
- Mitochondrial DNA Diversity of BroodstocDocument11 pagesMitochondrial DNA Diversity of Broodstocnazwar mashuriNo ratings yet
- Participatory Fishery Monitoring Is Successful For Understanding The Reproductive Biology Needed For Local Fisheries ManagementDocument16 pagesParticipatory Fishery Monitoring Is Successful For Understanding The Reproductive Biology Needed For Local Fisheries ManagementSelita AlTinNo ratings yet
- Discovery of The Invasive Mayan Cichlid Fish "Cichlasoma" UrophthalmusDocument18 pagesDiscovery of The Invasive Mayan Cichlid Fish "Cichlasoma" UrophthalmusViry BritoNo ratings yet
- Mirimin and Banguera Et Al 2011Document9 pagesMirimin and Banguera Et Al 2011IlonaNo ratings yet
- Alfaro Shigueto Et Al 2018 GillnetsDocument8 pagesAlfaro Shigueto Et Al 2018 GillnetsEventos ACNo ratings yet
- 2009 Molecular Systematics Taxonomy Biogeography Cavia - Dunnun - SalazarBravoDocument14 pages2009 Molecular Systematics Taxonomy Biogeography Cavia - Dunnun - SalazarBravoLucía HNo ratings yet
- Assessing Functions of Movement in A Great Plains Endemic FishDocument21 pagesAssessing Functions of Movement in A Great Plains Endemic FishgagenahNo ratings yet
- Molecular Phylogenetics and EvolutionDocument10 pagesMolecular Phylogenetics and EvolutionJulio César Chávez GalarzaNo ratings yet
- SpinK 2016 Filogenia y Diversif TortugasDocument13 pagesSpinK 2016 Filogenia y Diversif TortugasAlma AguilarNo ratings yet
- Olson and Galvan 2002Document20 pagesOlson and Galvan 2002Brandon Mitchell Benavente NavarreteNo ratings yet
- Research Article: Serum Biochemistry of Lumpy Skin Disease Virus-Infected CattleDocument6 pagesResearch Article: Serum Biochemistry of Lumpy Skin Disease Virus-Infected CattleYumeChanNo ratings yet
- Absen Kuliah Perdana Biokvet 1 TGL 3-8-21Document5 pagesAbsen Kuliah Perdana Biokvet 1 TGL 3-8-21YumeChanNo ratings yet
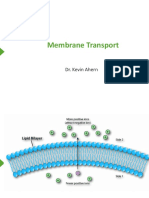
- Membrane Transport: Dr. Kevin AhernDocument32 pagesMembrane Transport: Dr. Kevin AhernYumeChanNo ratings yet
- Urticaria Pigmentosa Associated With Wilms TumorDocument3 pagesUrticaria Pigmentosa Associated With Wilms TumorYumeChanNo ratings yet
- L, T$WG: Ruriodur)Document4 pagesL, T$WG: Ruriodur)YumeChanNo ratings yet
- Cushing's Disease: A New Approach To Therapy in Equine and Canine PatientsDocument4 pagesCushing's Disease: A New Approach To Therapy in Equine and Canine PatientsYumeChanNo ratings yet
- Klabin Reports 2nd Quarter Earnings of R$ 15 Million: HighlightsDocument10 pagesKlabin Reports 2nd Quarter Earnings of R$ 15 Million: HighlightsKlabin_RINo ratings yet
- The Systems' Institute of Hindu ASTROLOGY, GURGAON (INDIA) (Registered)Document8 pagesThe Systems' Institute of Hindu ASTROLOGY, GURGAON (INDIA) (Registered)SiddharthSharmaNo ratings yet
- Describing LearnersDocument29 pagesDescribing LearnersSongül Kafa67% (3)
- Julie Jacko - Professor of Healthcare InformaticsDocument1 pageJulie Jacko - Professor of Healthcare InformaticsjuliejackoNo ratings yet
- Algebra. Equations. Solving Quadratic Equations B PDFDocument1 pageAlgebra. Equations. Solving Quadratic Equations B PDFRoberto CastroNo ratings yet
- Percy Bysshe ShelleyDocument20 pagesPercy Bysshe Shelleynishat_haider_2100% (1)
- Tugas Conditional Sentences YanneDocument3 pagesTugas Conditional Sentences Yanneyanne nurmalitaNo ratings yet
- Marketing PlanDocument41 pagesMarketing PlanMark AbainzaNo ratings yet
- Leg Res Cases 4Document97 pagesLeg Res Cases 4acheron_pNo ratings yet
- CEI and C4C Integration in 1602: Software Design DescriptionDocument44 pagesCEI and C4C Integration in 1602: Software Design Descriptionpkumar2288No ratings yet
- Writing - Hidden Curriculum v2 EditedDocument6 pagesWriting - Hidden Curriculum v2 EditedwhighfilNo ratings yet
- Activity Lesson PlanDocument2 pagesActivity Lesson PlanPsiho LoguseNo ratings yet
- Awareness and Usage of Internet Banking Facilities in Sri LankaDocument18 pagesAwareness and Usage of Internet Banking Facilities in Sri LankaTharindu Thathsarana RajapakshaNo ratings yet
- Bakhtin's Chronotope On The RoadDocument17 pagesBakhtin's Chronotope On The RoadLeandro OliveiraNo ratings yet
- The Role of Mahatma Gandhi in The Freedom Movement of IndiaDocument11 pagesThe Role of Mahatma Gandhi in The Freedom Movement of IndiaSwathi Prasad100% (6)
- Proper AdjectivesDocument3 pagesProper AdjectivesRania Mohammed0% (2)
- VtDA - The Ashen Cults (Vampire Dark Ages) PDFDocument94 pagesVtDA - The Ashen Cults (Vampire Dark Ages) PDFRafãoAraujo100% (1)
- #6 Decision Control InstructionDocument9 pages#6 Decision Control InstructionTimothy King LincolnNo ratings yet
- United States Court of Appeals, Third CircuitDocument1 pageUnited States Court of Appeals, Third CircuitScribd Government DocsNo ratings yet
- Sodium Borate: What Is Boron?Document2 pagesSodium Borate: What Is Boron?Gary WhiteNo ratings yet
- SassigbmentDocument7 pagesSassigbmentFurkaan Ali KhanNo ratings yet
- Theater 10 Syllabus Printed PDFDocument7 pagesTheater 10 Syllabus Printed PDFJim QuentinNo ratings yet
- Core ApiDocument27 pagesCore ApiAnderson Soares AraujoNo ratings yet
- MCN Drill AnswersDocument12 pagesMCN Drill AnswersHerne Balberde100% (1)
- A Quality Improvement Initiative To Engage Older Adults in The DiDocument128 pagesA Quality Improvement Initiative To Engage Older Adults in The Disara mohamedNo ratings yet
- Becoming FarmersDocument13 pagesBecoming FarmersJimena RoblesNo ratings yet
- Infusion Site Selection and Infusion Set ChangeDocument8 pagesInfusion Site Selection and Infusion Set ChangegaridanNo ratings yet
- Diode ExercisesDocument5 pagesDiode ExercisesbruhNo ratings yet
- What Is Folate WPS OfficeDocument4 pagesWhat Is Folate WPS OfficeMerly Grael LigligenNo ratings yet
- Nursing Care Plan: Pt.'s Data Nursing Diagnosis GoalsDocument1 pageNursing Care Plan: Pt.'s Data Nursing Diagnosis GoalsKiran Ali100% (3)