You might also like
- An Update On COVID-19 and Pregnancy: Djamieson@emory - EduDocument10 pagesAn Update On COVID-19 and Pregnancy: Djamieson@emory - EduRaul DoctoNo ratings yet
- 1998 Volvo s70-v70Document52 pages1998 Volvo s70-v70Raul DoctoNo ratings yet
- BMW 6 Cylinder Dual Vanos Piston Seal Repair Fix PDocument13 pagesBMW 6 Cylinder Dual Vanos Piston Seal Repair Fix PRaul DoctoNo ratings yet
- Satoru Takeda - New Assessment of Fetal Descent and Forceps Delivery (2018, Springer Singapore) PDFDocument44 pagesSatoru Takeda - New Assessment of Fetal Descent and Forceps Delivery (2018, Springer Singapore) PDFLuisNo ratings yet
- Seminar: Jan Claassen, Soojin ParkDocument17 pagesSeminar: Jan Claassen, Soojin ParkRaul DoctoNo ratings yet
- Central Venous Pressure (CVP)Document3 pagesCentral Venous Pressure (CVP)Raul DoctoNo ratings yet
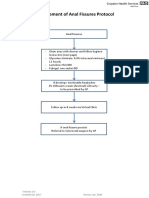
- Anal Fissures PDF 14 Sept-115042Document1 pageAnal Fissures PDF 14 Sept-115042Raul DoctoNo ratings yet
- GINA Coming SoonDocument1 pageGINA Coming SoonRaul DoctoNo ratings yet
- X3 E83 X3 3.0i FRNT Axle Support, Wishbone Tension StrutDocument3 pagesX3 E83 X3 3.0i FRNT Axle Support, Wishbone Tension StrutRaul DoctoNo ratings yet
- 1998 Volvo S70-V70Document52 pages1998 Volvo S70-V70cosminubNo ratings yet
- 2022 ACC A-Ha HFSAGuideline For The Management of Heart FailureDocument167 pages2022 ACC A-Ha HFSAGuideline For The Management of Heart FailureRaul DoctoNo ratings yet
- Ascoping Review of The Pathophysiology of COVID-19Document16 pagesAscoping Review of The Pathophysiology of COVID-19Raul DoctoNo ratings yet
- Rapid Diagnostic Testing For SARS-CoV-2Document9 pagesRapid Diagnostic Testing For SARS-CoV-2Wilfredo NuñezNo ratings yet
- Why Exercise Builds Muscles: Titin Mechanosensing Controls Skeletal Muscle Growth Under LoadDocument15 pagesWhy Exercise Builds Muscles: Titin Mechanosensing Controls Skeletal Muscle Growth Under LoadRaul Docto100% (1)
- Obstetrics & GynecologyDocument10 pagesObstetrics & GynecologyRaul DoctoNo ratings yet
- How To Install Google Play On The Amazon Fire HD 8Document7 pagesHow To Install Google Play On The Amazon Fire HD 8Raul DoctoNo ratings yet
- Robba2021 Article BasicUltrasoundHead-to-toeSkilDocument21 pagesRobba2021 Article BasicUltrasoundHead-to-toeSkilRaul DoctoNo ratings yet
- Complications of HysterectomyDocument17 pagesComplications of HysterectomyRaul DoctoNo ratings yet
- Standardization of Fetal Heart Rate Pattern Management: Is International Consensus Possible?Document8 pagesStandardization of Fetal Heart Rate Pattern Management: Is International Consensus Possible?Raul DoctoNo ratings yet
- AJOG Implementation of A Conservative ChecklistDocument9 pagesAJOG Implementation of A Conservative ChecklistRaul DoctoNo ratings yet
- Understanding The Role of Host Response in Influenza PneumonitisDocument3 pagesUnderstanding The Role of Host Response in Influenza PneumonitisRaul DoctoNo ratings yet
- Vlaar2021 Article TransfusionStrategiesInBleedinDocument25 pagesVlaar2021 Article TransfusionStrategiesInBleedinRaul DoctoNo ratings yet
- Case 35-2021: A 50-Year-Old Woman With Pain in The Left Upper Quadrant and HypoxemiaDocument7 pagesCase 35-2021: A 50-Year-Old Woman With Pain in The Left Upper Quadrant and HypoxemiaRaul DoctoNo ratings yet
- Alcohol y Metabolismo HumanoDocument21 pagesAlcohol y Metabolismo HumanoDani BelloNo ratings yet
- Surviving Sepsis Campaign International.21Document81 pagesSurviving Sepsis Campaign International.21jamesteryNo ratings yet
- Tugwell 1985Document13 pagesTugwell 1985Raul DoctoNo ratings yet
- A Framework For Standardized Management of Intrapartum PDFDocument6 pagesA Framework For Standardized Management of Intrapartum PDFRaul A UrbinaNo ratings yet
- 2021 - BMC - The - Japanese - Clinical - Practice - Guidelines - For - Management SepsisDocument144 pages2021 - BMC - The - Japanese - Clinical - Practice - Guidelines - For - Management SepsisSara MendesNo ratings yet
- MCU 2021 The Ventilator Book 3rd EditionDocument280 pagesMCU 2021 The Ventilator Book 3rd EditionAlvaro Estupiñan100% (2)
- 2014-08-13 Gynae Positioning Presentation - SlidesDocument14 pages2014-08-13 Gynae Positioning Presentation - SlidesIrina MincuNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Anatomy & Physiology of Female Reproductive SystemDocument5 pagesAnatomy & Physiology of Female Reproductive Systemaj_0852100% (2)
- Female Reproductive System AnatomyDocument21 pagesFemale Reproductive System AnatomyTrang KhúcNo ratings yet
- Endometriosis Dan Infertilitas: Alfaina WahyuniDocument15 pagesEndometriosis Dan Infertilitas: Alfaina WahyuniRudi GunawanNo ratings yet
- Recent Advancements in Infertility TreatmentsDocument71 pagesRecent Advancements in Infertility TreatmentsNimi SimonNo ratings yet
- Human SexualityDocument11 pagesHuman Sexualitylanilyn aquinoNo ratings yet
- Cowper's gland seminal fluid function reproductive anatomy menstrual cycle stagesDocument12 pagesCowper's gland seminal fluid function reproductive anatomy menstrual cycle stagescresia hidalgoNo ratings yet
- Intrauterine Drug Delivery SystemsDocument32 pagesIntrauterine Drug Delivery SystemsAhmad ThantowiNo ratings yet
- Hormonal Advances OlufemiDocument90 pagesHormonal Advances OlufemiDare Olalekan AnthonyNo ratings yet
- Hormone Replacement Therapy: Memorable Medicine Medical Revision Notes, Tutorials & ForumsDocument9 pagesHormone Replacement Therapy: Memorable Medicine Medical Revision Notes, Tutorials & ForumsMemMed AdministratorNo ratings yet
- Obstetrics NotesDocument114 pagesObstetrics NotesKiara Luna100% (2)
- 2.1 One Pager Female Reproduction MEMODocument1 page2.1 One Pager Female Reproduction MEMORudzi UdziNo ratings yet
- Pourusha VastiDocument54 pagesPourusha Vastiksr prasadNo ratings yet
- Clinical Use of Progestins in Bitches and Queens - A Review - IVISDocument35 pagesClinical Use of Progestins in Bitches and Queens - A Review - IVISDai AnaNo ratings yet
- (Midwifery Final) Pages 15 26Document12 pages(Midwifery Final) Pages 15 26AdhityaEkoBagusNo ratings yet
- Gynaecological Ultrasound in Clinical PracticeDocument247 pagesGynaecological Ultrasound in Clinical Practiceyulb_1100% (5)
- MORNING REPORT OBSTETRICS AND GYNECOLOGYDocument16 pagesMORNING REPORT OBSTETRICS AND GYNECOLOGYSiti NofriansyahNo ratings yet
- The Male UrethraDocument5 pagesThe Male UrethraajescoolNo ratings yet
- Human Reproduction Module 1 - Development and Function of Reproductive SystemDocument55 pagesHuman Reproduction Module 1 - Development and Function of Reproductive SystemAdc ClamorNo ratings yet
- Pathogenesis of PcosDocument14 pagesPathogenesis of PcosMitali Narurkar100% (1)
- Family Planning Methods for a Healthy FamilyDocument36 pagesFamily Planning Methods for a Healthy FamilyLive with CjNo ratings yet
- (A) Zygote: (C) Fish 3Document2 pages(A) Zygote: (C) Fish 3UGC GAMERNo ratings yet
- Sex Determination in MammalsDocument33 pagesSex Determination in MammalsBinita SedhaiNo ratings yet
- Name: Class:: Natural Science Primary 6 - Downloadable and Printable © Ediciones Bilingües, S.LDocument2 pagesName: Class:: Natural Science Primary 6 - Downloadable and Printable © Ediciones Bilingües, S.LMªDolores86% (14)
- Lyphochek Immunoassay Plus Control Levels 1, 2 and 3Document1 pageLyphochek Immunoassay Plus Control Levels 1, 2 and 3aa shuvoNo ratings yet
- Basic Biology of Sexual BehaviorDocument4 pagesBasic Biology of Sexual BehaviornegriRDNo ratings yet
- Abnormal Uterine Bleeding - PresentationDocument76 pagesAbnormal Uterine Bleeding - PresentationlookNo ratings yet
- Gynecologic NursingDocument218 pagesGynecologic Nursingblacklilha100% (1)
- Anatomy and Physiology of ReproductionDocument53 pagesAnatomy and Physiology of ReproductionPrincess Diane M. RagusNo ratings yet
- Na Blog - CZ: Reproduction ClozeDocument6 pagesNa Blog - CZ: Reproduction ClozeUtari Septiana DewiNo ratings yet
- Primary Amenorrhoea 2Document42 pagesPrimary Amenorrhoea 2sivaNo ratings yet